

Ultrahigh-throughput discovery of promiscuous enzymes by picodroplet functional metagenomics
- PMID: 26639611
- PMCID: PMC4686663
- DOI: 10.1038/ncomms10008
Ultrahigh-throughput discovery of promiscuous enzymes by picodroplet functional metagenomics
Abstract
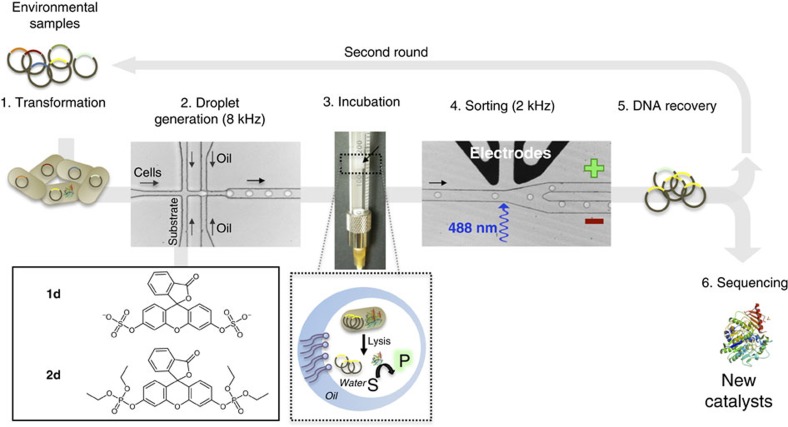
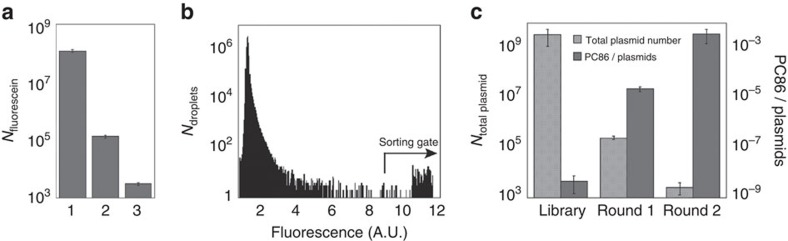
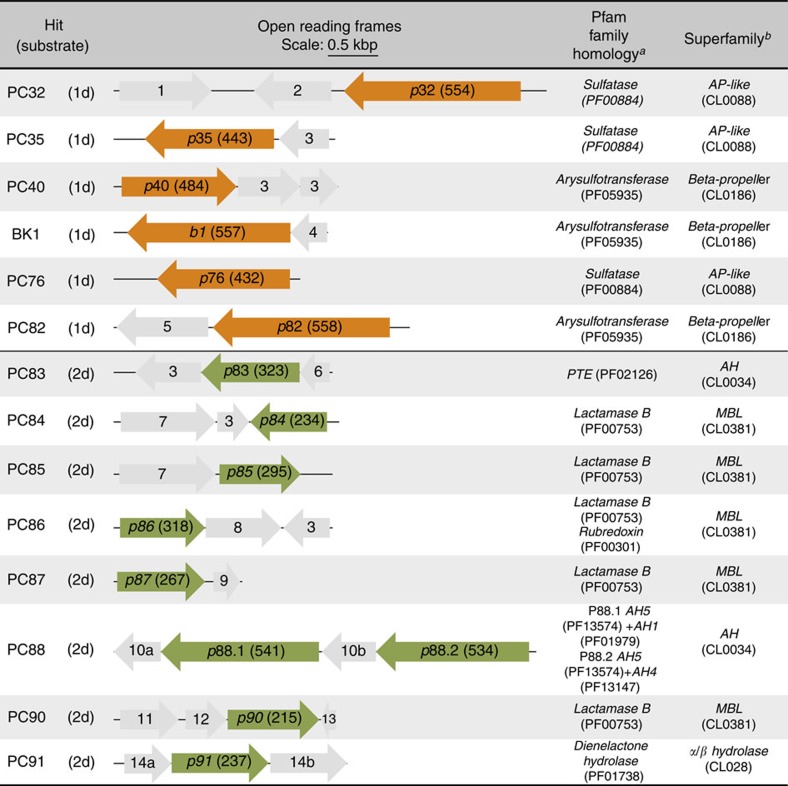
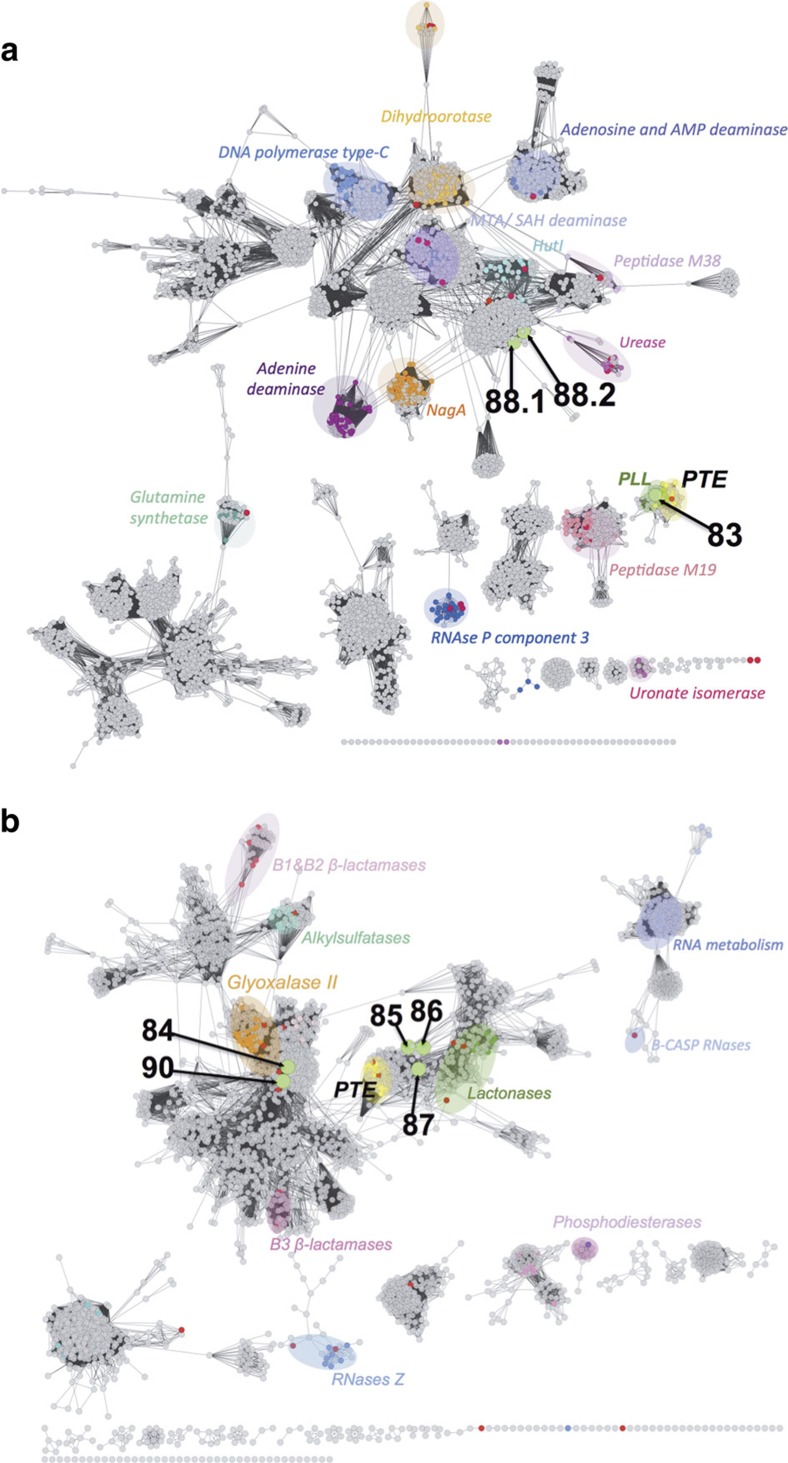
Unculturable bacterial communities provide a rich source of biocatalysts, but their experimental discovery by functional metagenomics is difficult, because the odds are stacked against the experimentor. Here we demonstrate functional screening of a million-membered metagenomic library in microfluidic picolitre droplet compartments. Using bait substrates, new hydrolases for sulfate monoesters and phosphotriesters were identified, mostly based on promiscuous activities presumed not to be under selection pressure. Spanning three protein superfamilies, these break new ground in sequence space: promiscuity now connects enzymes with only distantly related sequences. Most hits could not have been predicted by sequence analysis, because the desired activities have never been ascribed to similar sequences, showing how this approach complements bioinformatic harvesting of metagenomic sequencing data. Functional screening of a library of unprecedented size with excellent assay sensitivity has been instrumental in identifying rare genes constituting catalytically versatile hubs in sequence space as potential starting points for the acquisition of new functions.
Figures
References
-
- Bornscheuer U. T. et al. Engineering the third wave of biocatalysis. Nature 485, 185–194 (2012). - PubMed
-
- Daniel R. The metagenomics of soil. Nat. Rev. Microbiol. 3, 470–478 (2005). - PubMed
-
- Venter J. C. et al. Environmental genome shotgun sequencing of the Sargasso Sea. Science 304, 66–74 (2004). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
