

Injured sensory neuron-derived CSF1 induces microglial proliferation and DAP12-dependent pain
- PMID: 26642091
- PMCID: PMC4703328
- DOI: 10.1038/nn.4189
Injured sensory neuron-derived CSF1 induces microglial proliferation and DAP12-dependent pain
Abstract
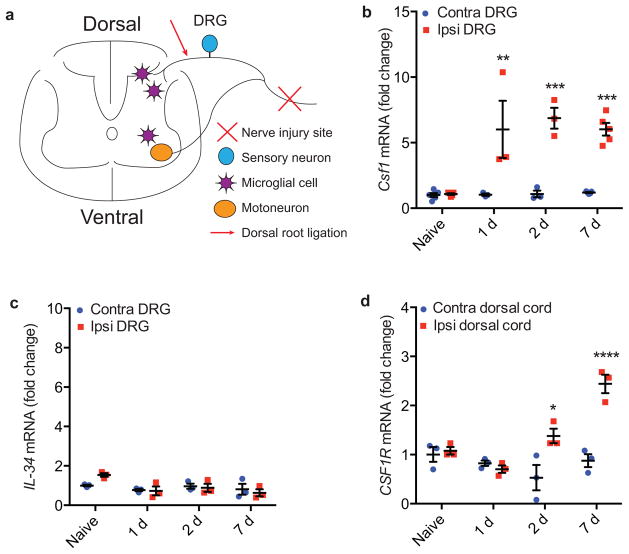
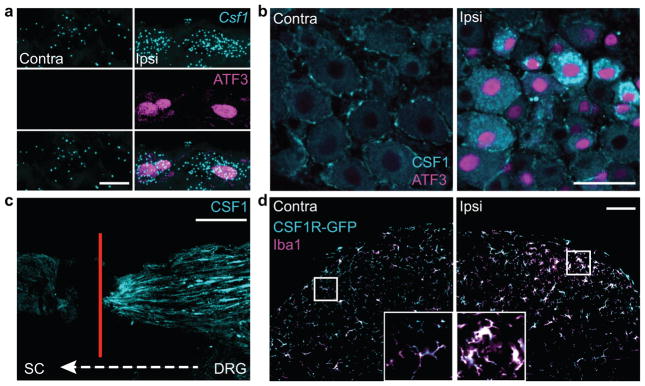
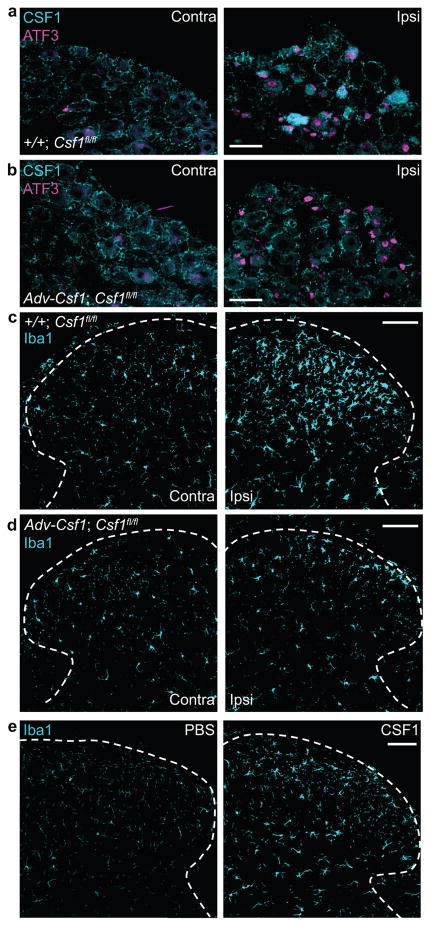
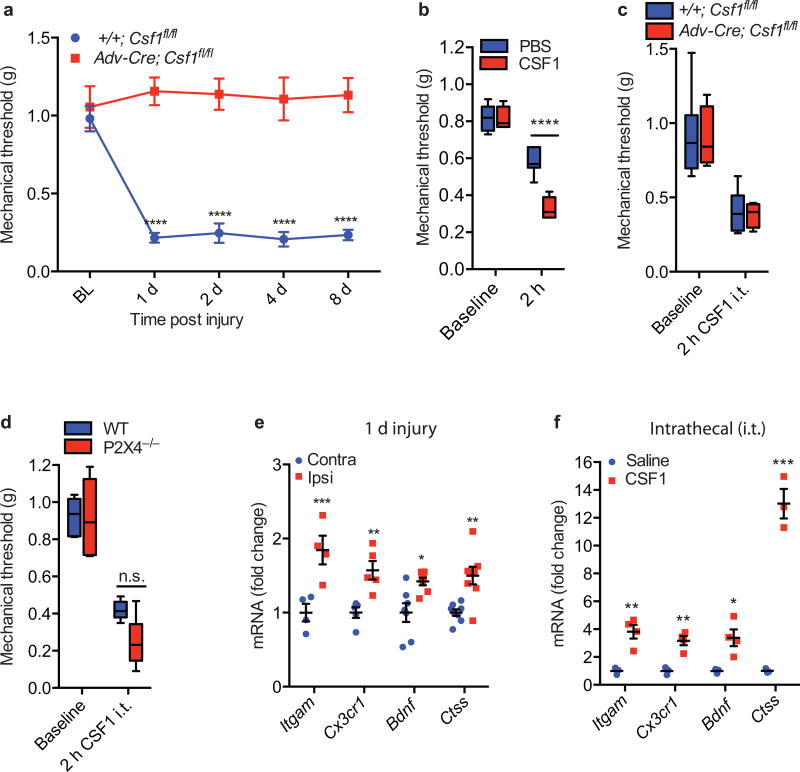
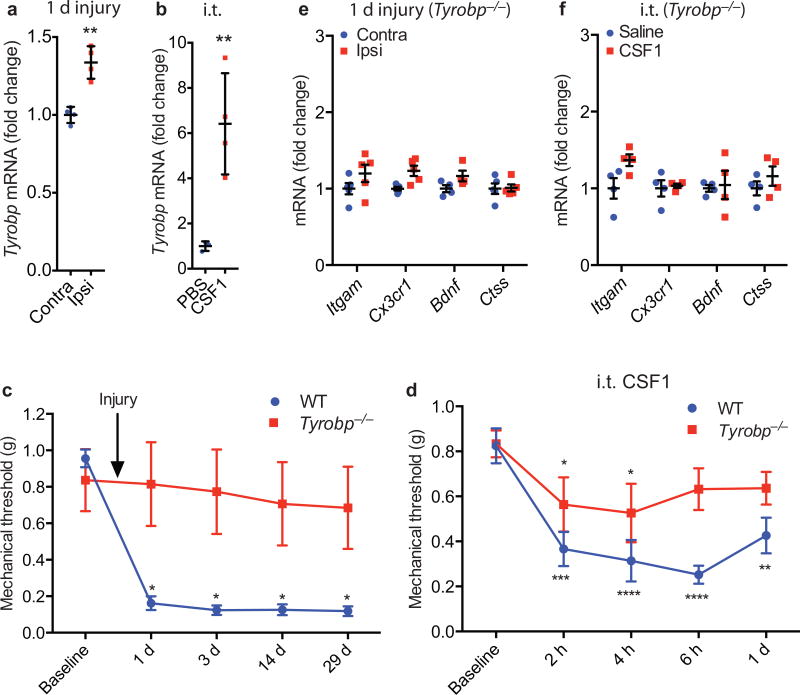
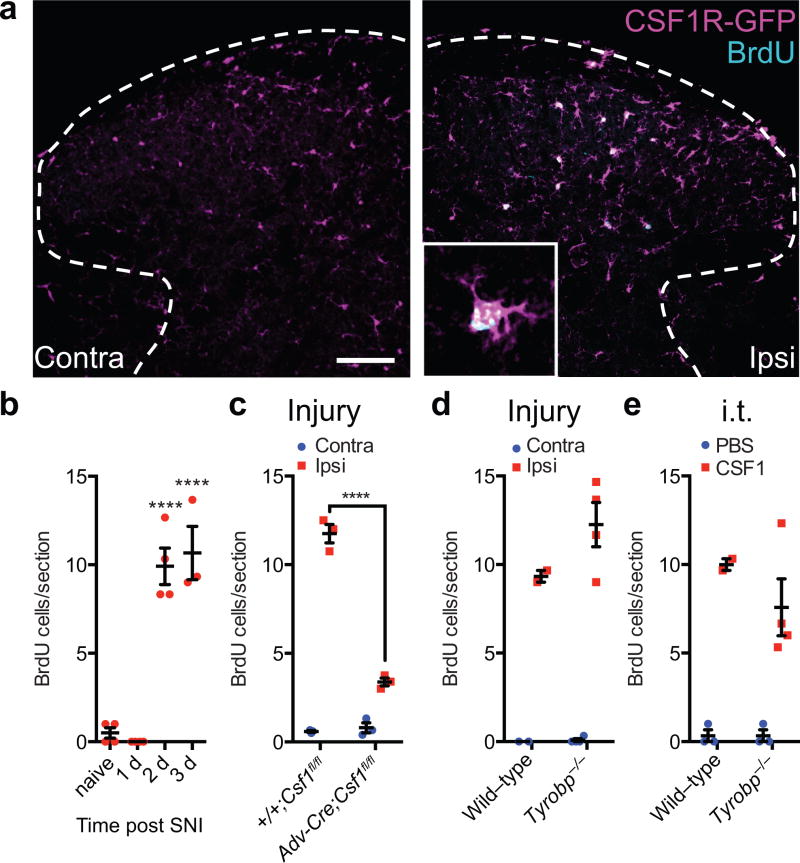
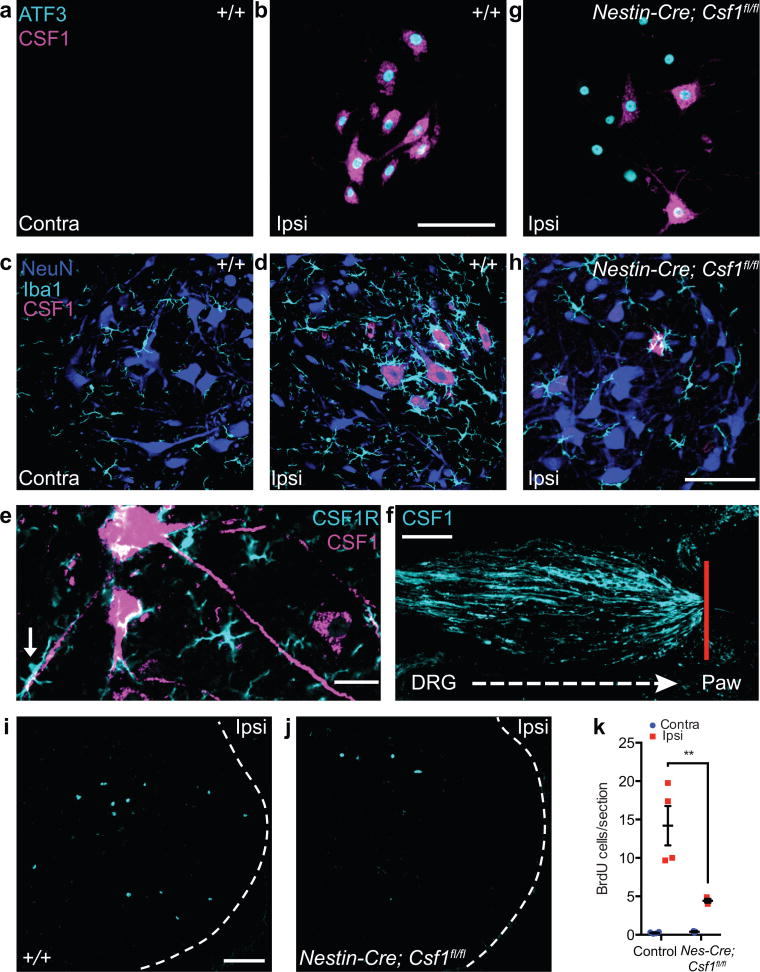
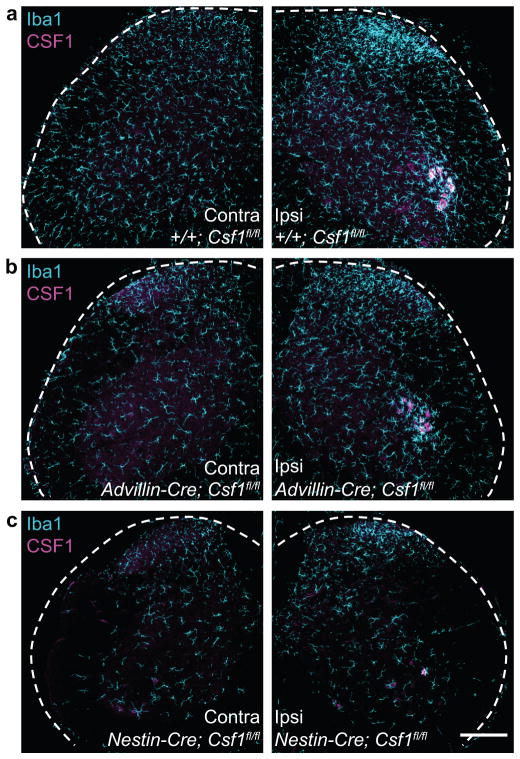
Although microglia have been implicated in nerve injury-induced neuropathic pain, the manner by which injured sensory neurons engage microglia remains unclear. We found that peripheral nerve injury induced de novo expression of colony-stimulating factor 1 (CSF1) in injured sensory neurons. CSF1 was transported to the spinal cord, where it targeted the microglial CSF1 receptor (CSF1R). Cre-mediated sensory neuron deletion of Csf1 completely prevented nerve injury-induced mechanical hypersensitivity and reduced microglial activation and proliferation. In contrast, intrathecal injection of CSF1 induced mechanical hypersensitivity and microglial proliferation. Nerve injury also upregulated CSF1 in motoneurons, where it was required for ventral horn microglial activation and proliferation. Downstream of CSF1R, we found that the microglial membrane adaptor protein DAP12 was required for both nerve injury- and intrathecal CSF1-induced upregulation of pain-related microglial genes and the ensuing pain, but not for microglial proliferation. Thus, both CSF1 and DAP12 are potential targets for the pharmacotherapy of neuropathic pain.
Figures
Comment in
-
A peripheral messenger for chronic pain.Nat Neurosci. 2016 Jan;19(1):9. doi: 10.1038/nn.4217. Nat Neurosci. 2016. PMID: 26713743 No abstract available.
References
-
- Backonja M, Woolf CJ. Future directions in neuropathic pain therapy: closing the translational loop. Oncologist. 2010;15(Suppl 2):24–29. - PubMed
-
- Sieweke MH, Allen JE. Beyond stem cells: self-renewal of differentiated macrophages. Science. 2013;342:1242974. - PubMed
-
- Salter MW, Beggs S. Sublime microglia: expanding roles for the guardians of the CNS. Cell. 2014;158:15–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AG045040/AG/NIA NIH HHS/United States
- DE022001/DE/NIDCR NIH HHS/United States
- K08 NS078050/NS/NINDS NIH HHS/United States
- AI068129/AI/NIAID NIH HHS/United States
- R01 NS014627/NS/NINDS NIH HHS/United States
- NS14627/NS/NINDS NIH HHS/United States
- K08NS078050/NS/NINDS NIH HHS/United States
- R37 NS014627/NS/NINDS NIH HHS/United States
- R01 AG045040/AG/NIA NIH HHS/United States
- R01 AI068129/AI/NIAID NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 DA036985/DA/NIDA NIH HHS/United States
- R01 DE022001/DE/NIDCR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
