

Gates, Channels, and Switches: Elements of the Proteasome Machine
- PMID: 26643069
- PMCID: PMC4706478
- DOI: 10.1016/j.tibs.2015.10.009
Gates, Channels, and Switches: Elements of the Proteasome Machine
Abstract
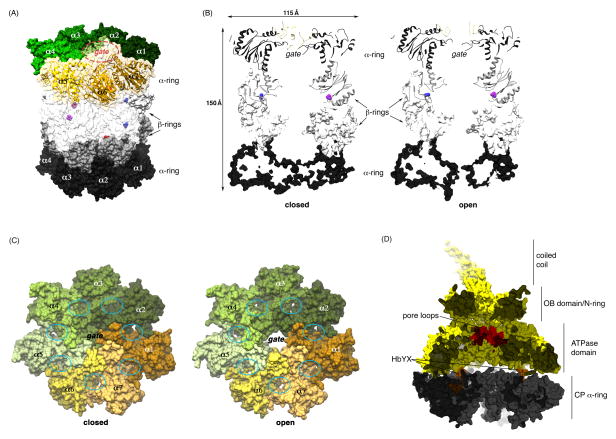
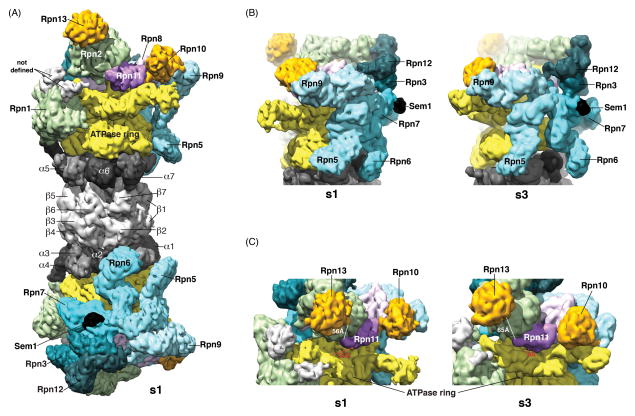
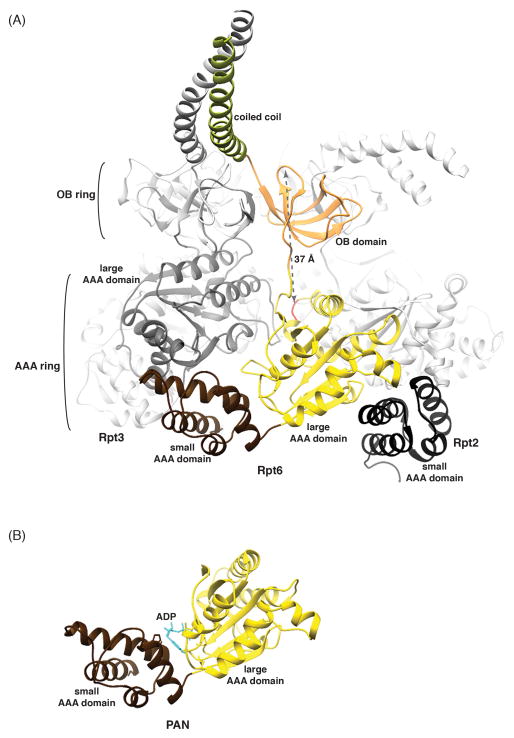
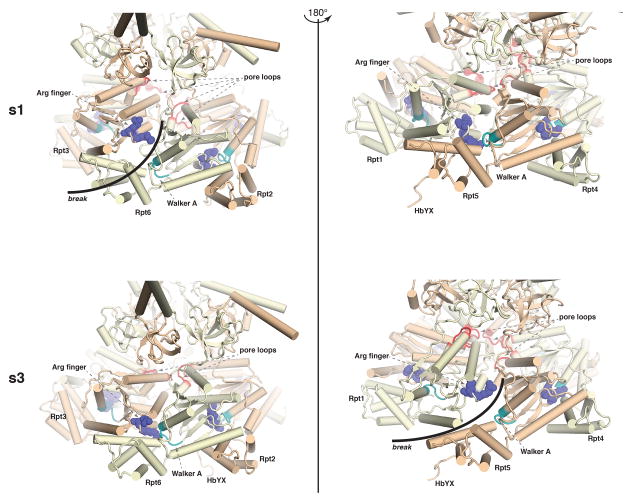
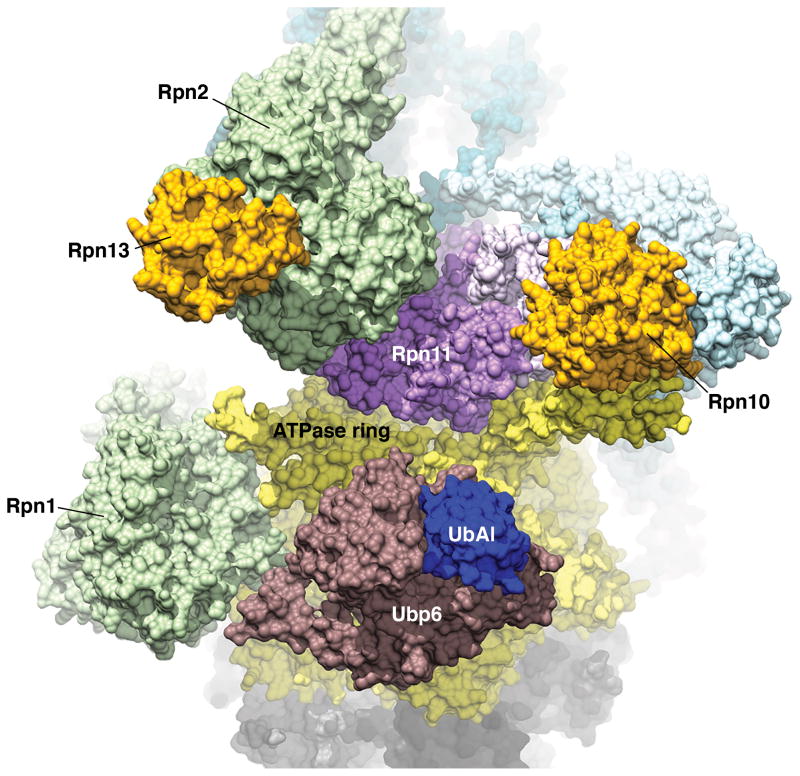
The proteasome has emerged as an intricate machine that has dynamic mechanisms to regulate the timing of its activity, its selection of substrates, and its processivity. The 19-subunit regulatory particle (RP) recognizes ubiquitinated proteins, removes ubiquitin, and injects the target protein into the proteolytic chamber of the core particle (CP) via a narrow channel. Translocation into the CP requires substrate unfolding, which is achieved through mechanical force applied by a hexameric ATPase ring of the RP. Recent cryoelectron microscopy (cryoEM) studies have defined distinct conformational states of the RP, providing illustrative snapshots of what appear to be progressive steps of substrate engagement. Here, we bring together this new information with molecular analyses to describe the principles of proteasome activity and regulation.
Keywords: ATPase; proteasome; protein degradation; ubiquitin.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Groll M, et al. Structure of 20S proteasome from yeast at 2.4 A resolution. Nature. 1997;386:463–471. - PubMed
-
- Whitby FG, et al. Structural basis for the activation of 20S proteasomes by 11S regulators. Nature. 2000;408:115–120. - PubMed
-
- Groll M, et al. A gated channel into the proteasome core particle. Nat Struct Biol. 2000;7:1062–1067. - PubMed
-
- Weber-Ban EU, Reid BG, Miranker AD, Horwich AL. Global unfolding of a substrate protein by the Hsp100 chaperone ClpA. Nature. 1999;401:90–93. - PubMed
-
- Lee C, Schwartz MP, Prakash S, Iwakura M, Matouschek A. ATP-dependent proteases degrade their substrates by processively unraveling them from the degradation signal. Mol Cell. 2001;7:627–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
