

Naturally Occurring Subclinical Endotoxemia in Humans Alters Adaptive and Innate Immune Functions through Reduced MAPK and Increased STAT1 Phosphorylation
- PMID: 26643479
- PMCID: PMC4707069
- DOI: 10.4049/jimmunol.1501888
Naturally Occurring Subclinical Endotoxemia in Humans Alters Adaptive and Innate Immune Functions through Reduced MAPK and Increased STAT1 Phosphorylation
Abstract
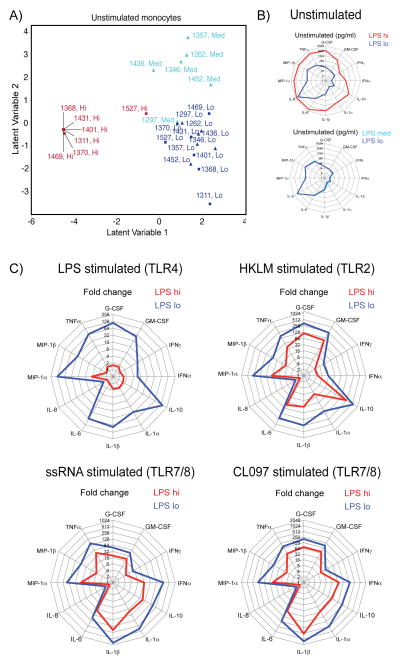
Multiple studies have shown correlates of immune activation with microbial translocation and plasma LPS during HIV infection. It is unclear whether this activation is due to LPS, residual viral replication, or both. Few studies have addressed the effects of persistent in vivo levels of LPS on specific immune functions in humans in the absence of chronic viral infection or pathological settings such as sepsis. We previously reported on a cohort of HIV-negative men with subclinical endotoxemia linked to alterations in CD4/CD8 T cell ratio and plasma cytokine levels. This HIV-negative cohort allowed us to assess cellular immune functions in the context of different subclinical plasma LPS levels ex vivo without confounding viral effects. By comparing two samples of differing plasma LPS levels from each individual, we now show that subclinical levels of plasma LPS in vivo significantly alter T cell proliferative capacity, monocyte cytokine release, and HLA-DR expression, and induce TLR cross-tolerance by decreased phosphorylation of MAPK pathway components. Using this human in vivo model of subclinical endotoxemia, we furthermore show that plasma LPS leads to constitutive activation of STAT1 through autocrine cytokine signaling, suggesting that subclinical endotoxemia in healthy individuals might lead to significant changes in immune function that have thus far not been appreciated.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures





References
-
- Caradonna L, Amati L, Magrone T, Pellegrino NM, Jirillo E, Caccavo D. Enteric bacteria, lipopolysaccharides and related cytokines in inflammatory bowel disease: biological and clinical significance. Journal of endotoxin research. 2000;6:205–214. - PubMed
-
- Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Bornstein E, Lambotte O, Altmann D, Blazar BR, Rodriguez B, Teixeira-Johnson L, Landay A, Martin JN, Hecht FM, Picker LJ, Lederman MM, Deeks SG, Douek DC. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nature medicine. 2006;12:1365–1371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
