

Thyroid hormone and vitamin D regulate VGF expression and promoter activity
- PMID: 26643910
- PMCID: PMC4705542
- DOI: 10.1530/JME-15-0224
Thyroid hormone and vitamin D regulate VGF expression and promoter activity
Abstract
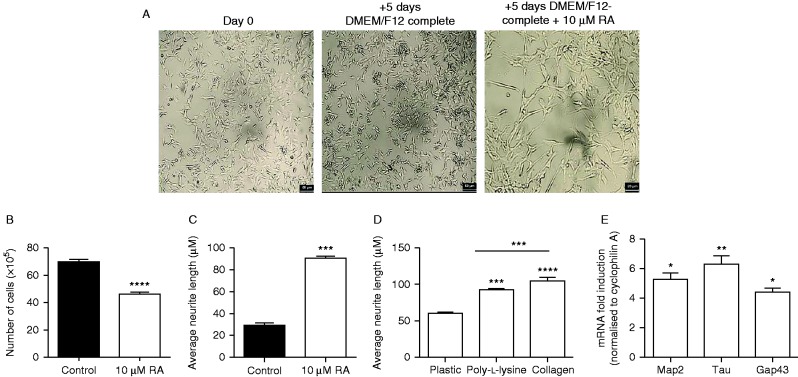
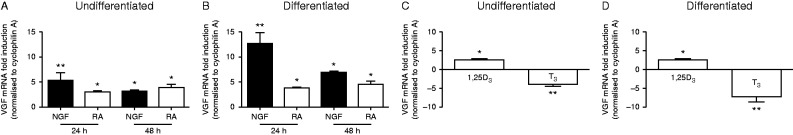
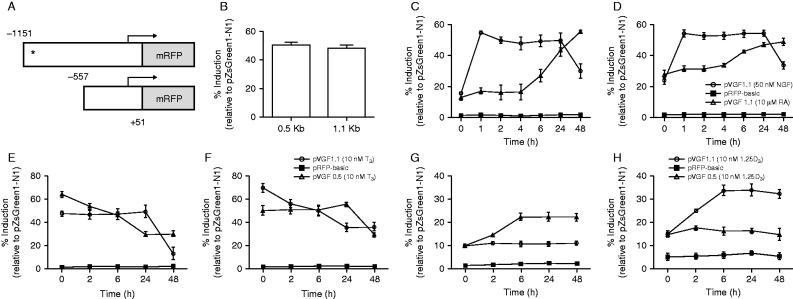
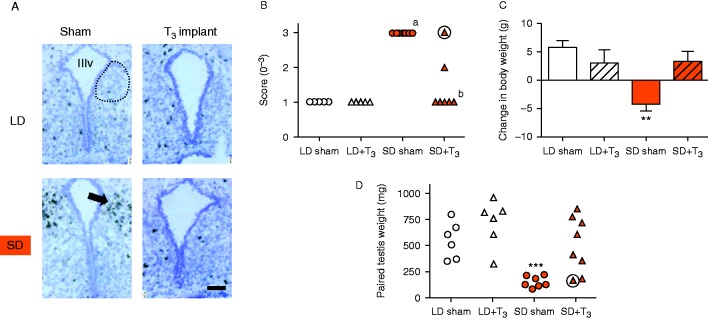
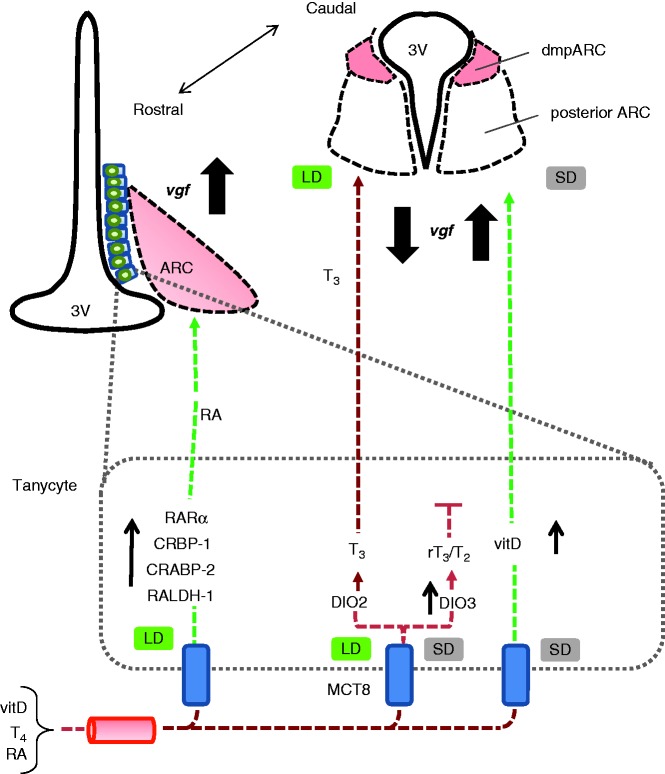
The Siberian hamster (Phodopus sungorus) survives winter by decreasing food intake and catabolizing abdominal fat reserves, resulting in a sustained, profound loss of body weight. Hypothalamic tanycytes are pivotal for this process. In these cells, short-winter photoperiods upregulate deiodinase 3, an enzyme that regulates thyroid hormone availability, and downregulate genes encoding components of retinoic acid (RA) uptake and signaling. The aim of the current studies was to identify mechanisms by which seasonal changes in thyroid hormone and RA signaling from tanycytes might ultimately regulate appetite and energy expenditure. proVGF is one of the most abundant peptides in the mammalian brain, and studies have suggested a role for VGF-derived peptides in the photoperiodic regulation of body weight in the Siberian hamster. In silico studies identified possible thyroid and vitamin D response elements in the VGF promoter. Using the human neuroblastoma SH-SY5Y cell line, we demonstrate that RA increases endogenous VGF expression (P<0.05) and VGF promoter activity (P<0.0001). Similarly, treatment with 1,25-dihydroxyvitamin D3 increased endogenous VGF mRNA expression (P<0.05) and VGF promoter activity (P<0.0001), whereas triiodothyronine (T3) decreased both (P<0.01 and P<0.0001). Finally, intra-hypothalamic administration of T3 blocked the short day-induced increase in VGF expression in the dorsomedial posterior arcuate nucleus of Siberian hamsters. Thus, we conclude that VGF expression is a likely target of photoperiod-induced changes in tanycyte-derived signals and is potentially a regulator of seasonal changes in appetite and energy expenditure.
Keywords: SH-SY5Y cells; Siberian hamster; VGF (non-acronymic); thyroid hormone; vitamin D.
© 2016 The authors.
Figures
References
-
- Agholme L, Lindstrom T, Kagedal K, Marcusson J, Hallbeck M. An in vitro model for neuroscience: differentiation of SH-SY5Y cells into cells with morphological and biochemical characteristics of mature neurons. Journal of Alzheimer's Disease. 2010;20:1069–1082. doi: 10.3233/JAD-2010-091363. - DOI - PubMed
-
- Anzano MA, Lamb AJ, Olson JA. Growth, appetite, sequence of pathological signs and survival following the induction of rapid, synchronous vitamin A deficiency in the rat. Journal of Nutrition. 1979;109:1419–1431. - PubMed
-
- Barrett P, Ross AW, Balik A, Littlewood PA, Mercer JG, Moar KM, Sallmen T, Kaslin J, Panula P, Schuhler S, et al. Photoperiodic regulation of histamine H3 receptor and VGF messenger ribonucleic acid in the arcuate nucleus of the Siberian hamster. Endocrinology. 2005;146:1930–1939. doi: 10.1210/en.2004-1452. - DOI - PubMed
-
- Barrett P, Ivanova E, Graham ES, Ross AW, Wilson D, Ple H, Mercer JG, Ebling FJ, Schuhler S, Dupre SM, et al. Photoperiodic regulation of cellular retinol binding protein, CRBP1 [corrected] and nestin in tanycytes of the third ventricle ependymal layer of the Siberian hamster. Journal of Endocrinology. 2006;191:687–698. doi: 10.1677/joe.1.06929. - DOI - PubMed
-
- Barrett P, Ebling FJ, Schuhler S, Wilson D, Ross AW, Warner A, Jethwa P, Boelen A, Visser TJ, Ozanne DM, et al. Hypothalamic thyroid hormone catabolism acts as a gatekeeper for the seasonal control of body weight and reproduction. Endocrinology. 2007;148:3608–3617. doi: 10.1210/en.2007-0316. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
