

Ephrin-A3 promotes and maintains slow muscle fiber identity during postnatal development and reinnervation
- PMID: 26644518
- PMCID: PMC4674275
- DOI: 10.1083/jcb.201502036
Ephrin-A3 promotes and maintains slow muscle fiber identity during postnatal development and reinnervation
Abstract
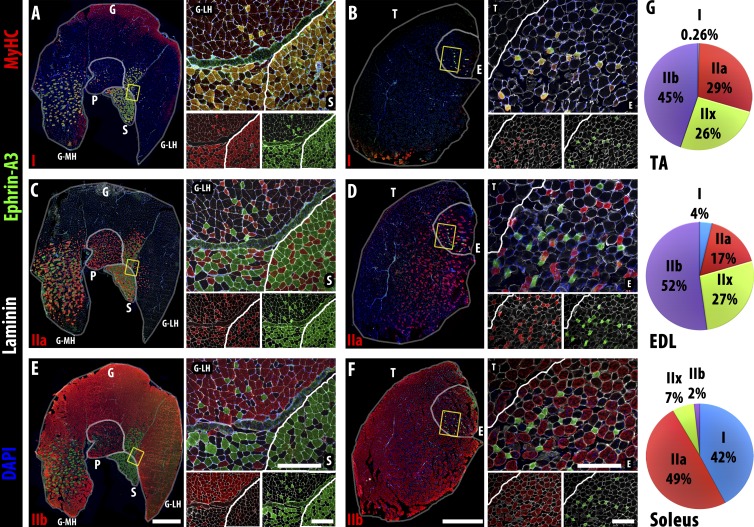
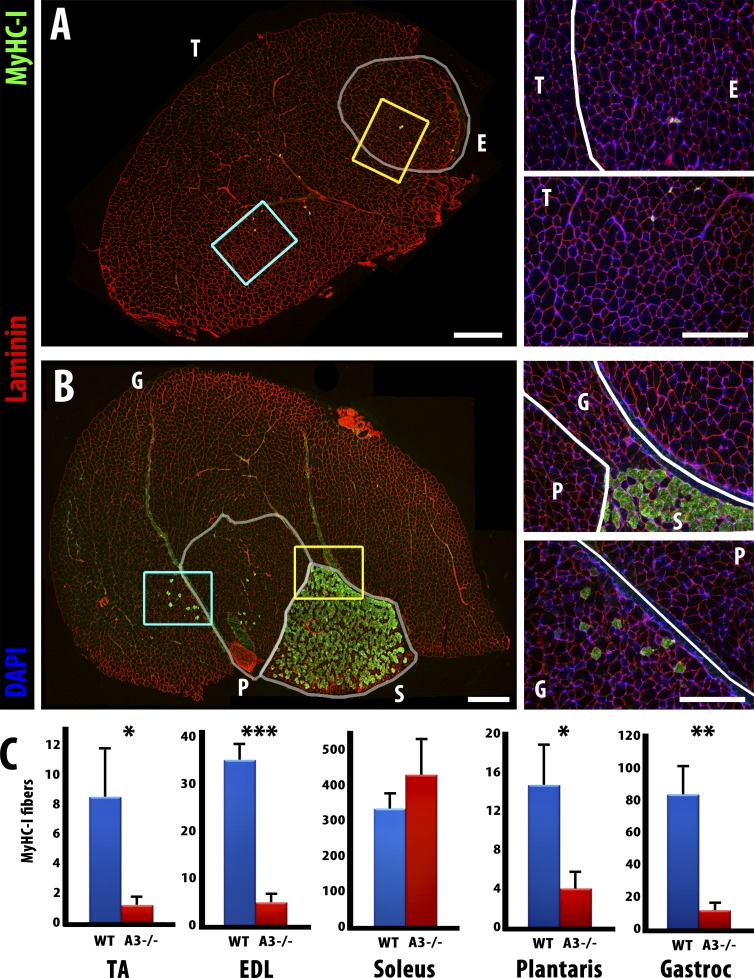
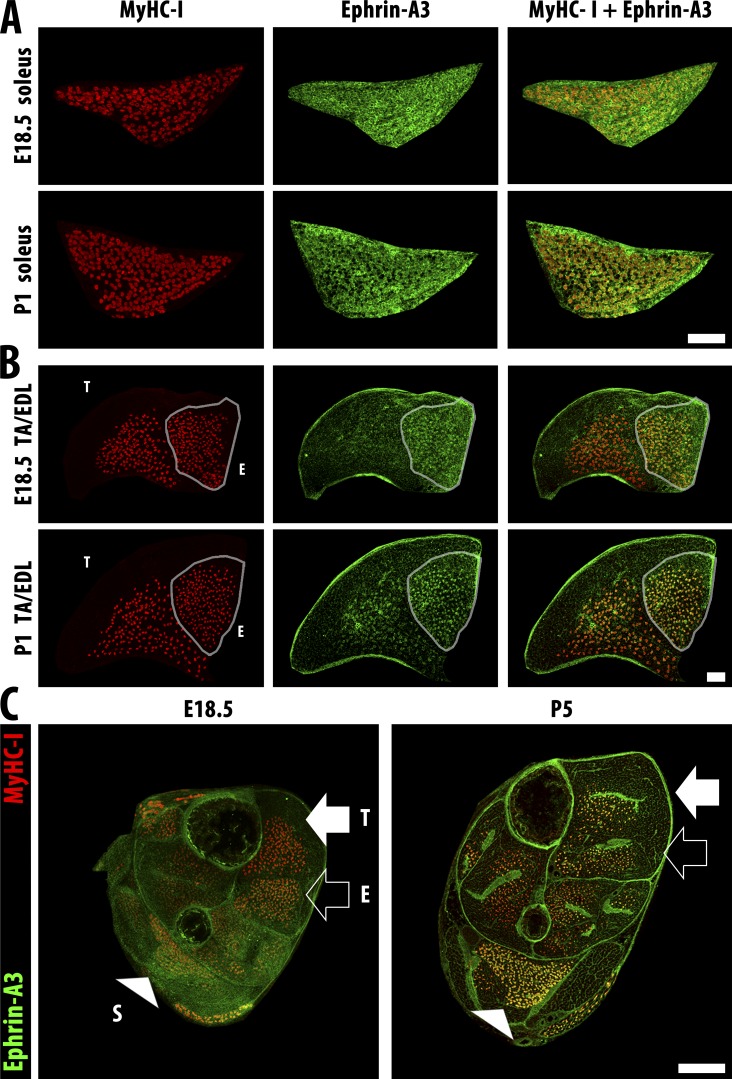
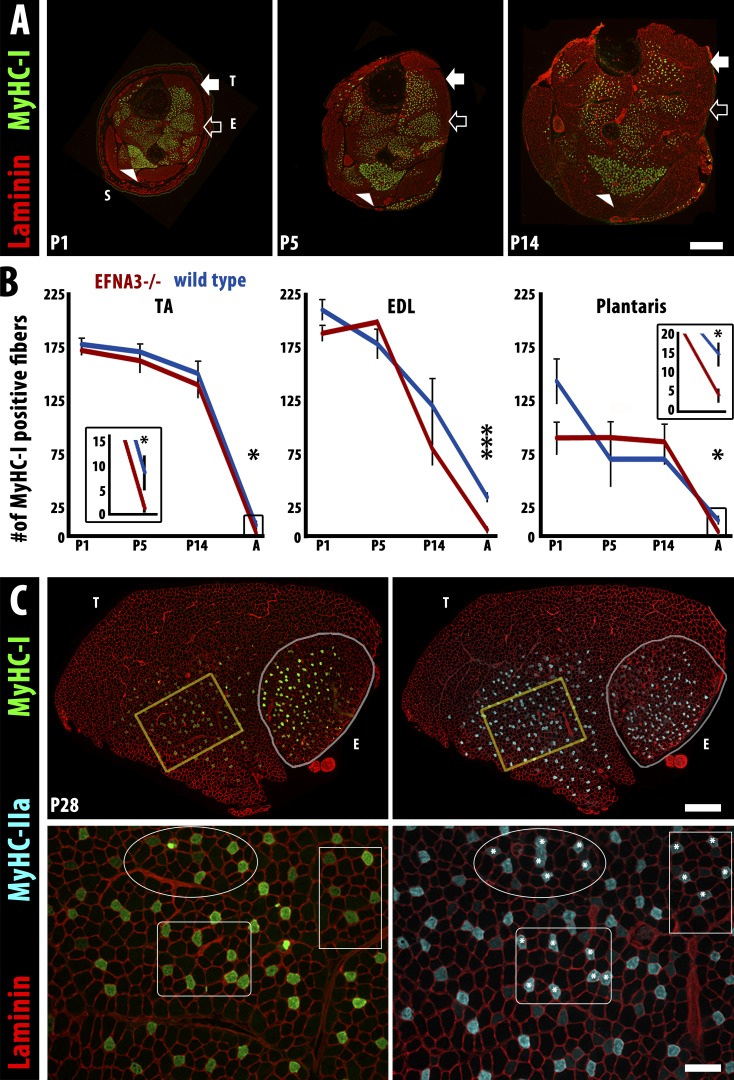
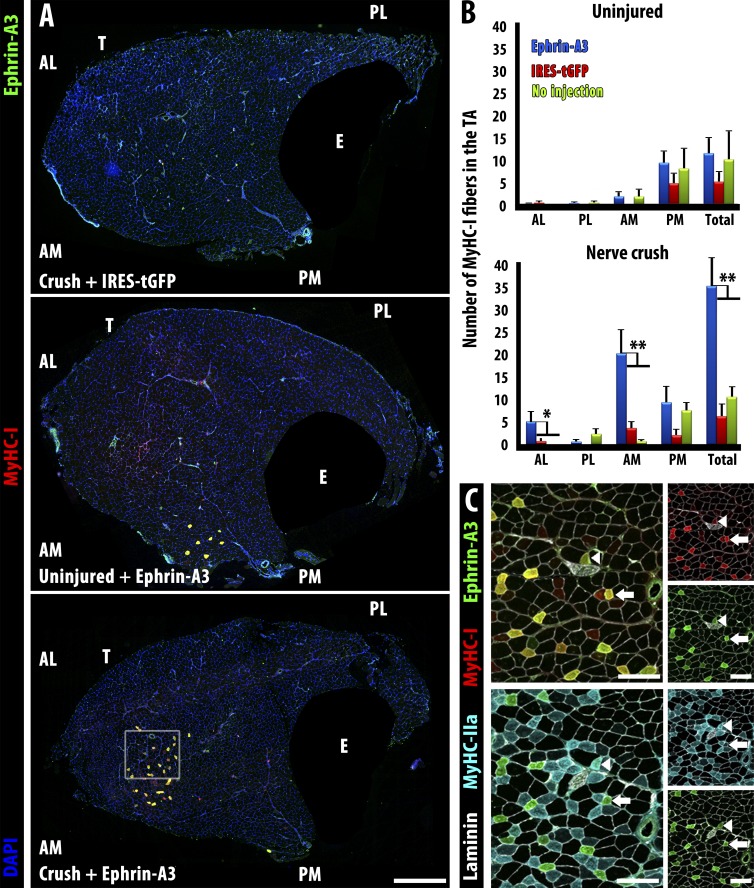
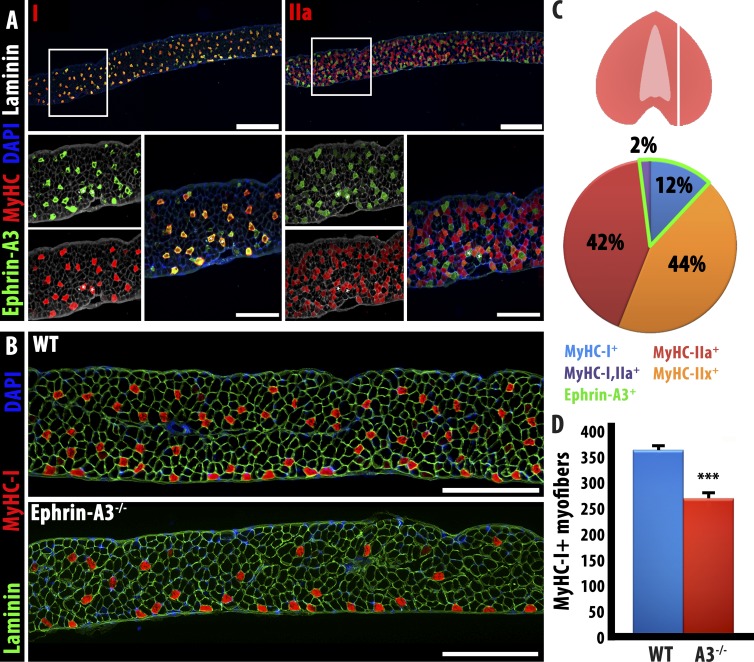
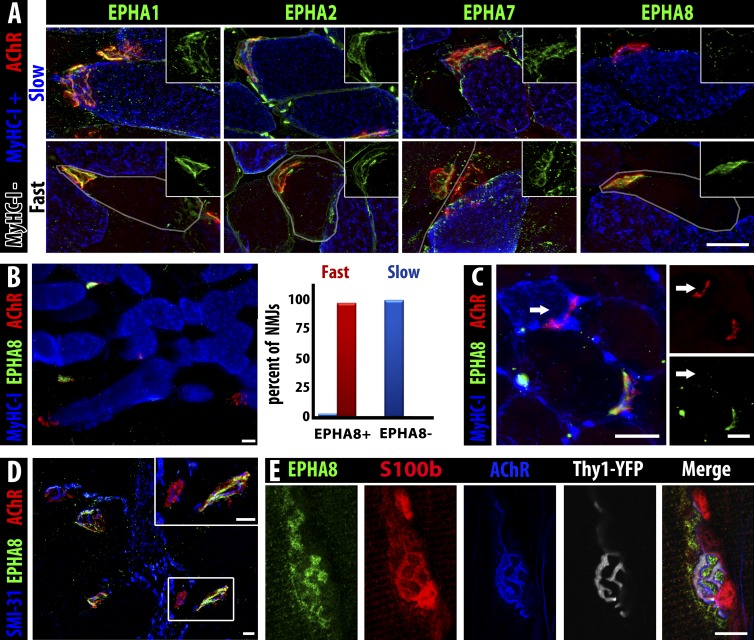
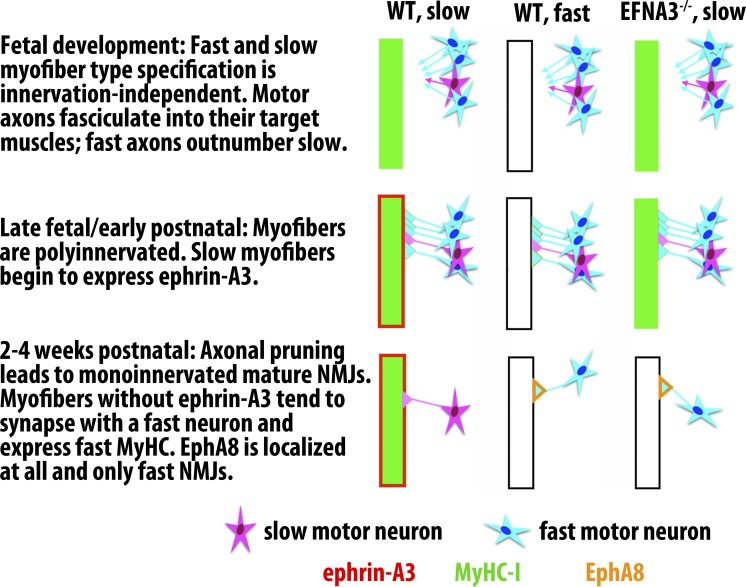
Each adult mammalian skeletal muscle has a unique complement of fast and slow myofibers, reflecting patterns established during development and reinforced via their innervation by fast and slow motor neurons. Existing data support a model of postnatal "matching" whereby predetermined myofiber type identity promotes pruning of inappropriate motor axons, but no molecular mechanism has yet been identified. We present evidence that fiber type-specific repulsive interactions inhibit innervation of slow myofibers by fast motor axons during both postnatal maturation of the neuromuscular junction and myofiber reinnervation after injury. The repulsive guidance ligand ephrin-A3 is expressed only on slow myofibers, whereas its candidate receptor, EphA8, localizes exclusively to fast motor endplates. Adult mice lacking ephrin-A3 have dramatically fewer slow myofibers in fast and mixed muscles, and misexpression of ephrin-A3 on fast myofibers followed by denervation/reinnervation promotes their respecification to a slow phenotype. We therefore conclude that Eph/ephrin interactions guide the fiber type specificity of neuromuscular interactions during development and adult life.
© 2015 Stark et al.
Figures
References
-
- Augusto V., Padovani C.R., and Campos G.E.R.. 2004. Skeletal muscle fiber types in C57BL6 mice. Braz. J. Morphol. Sci. 21:89–94.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
