

Capturing complex tumour biology in vitro: histological and molecular characterisation of precision cut slices
- PMID: 26647838
- PMCID: PMC4673528
- DOI: 10.1038/srep17187
Capturing complex tumour biology in vitro: histological and molecular characterisation of precision cut slices
Abstract
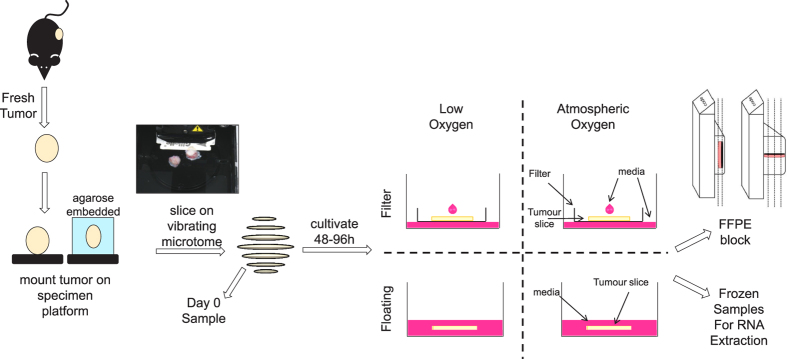
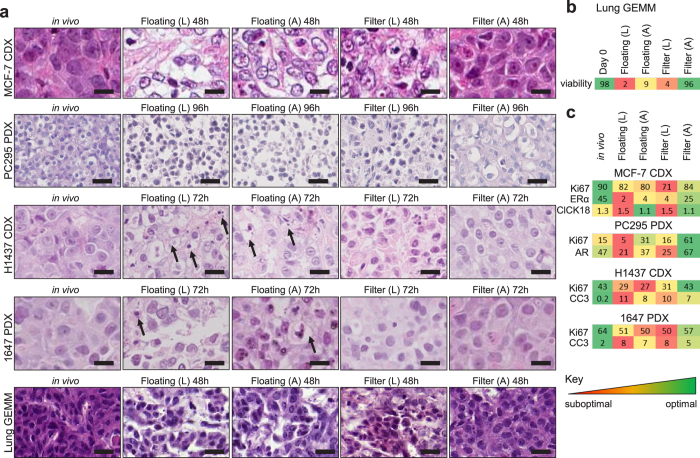
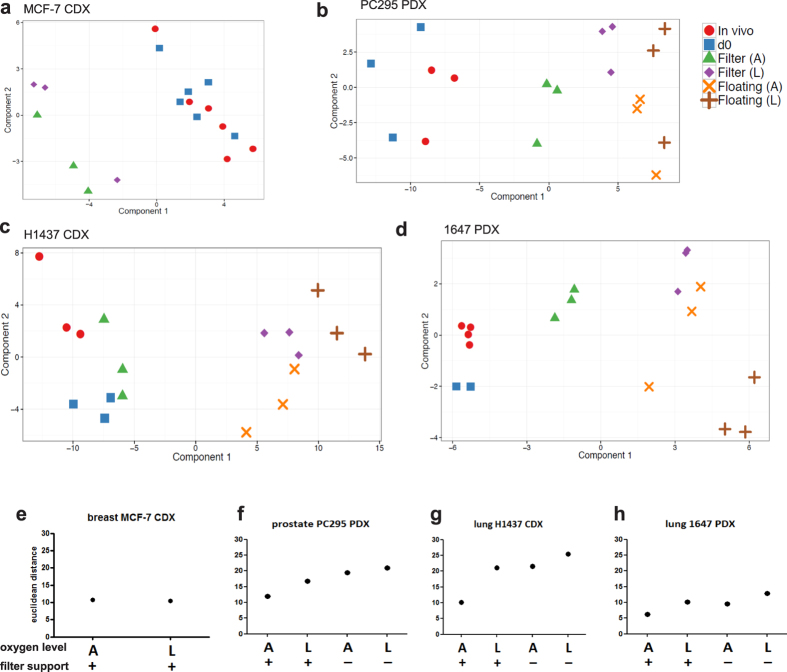
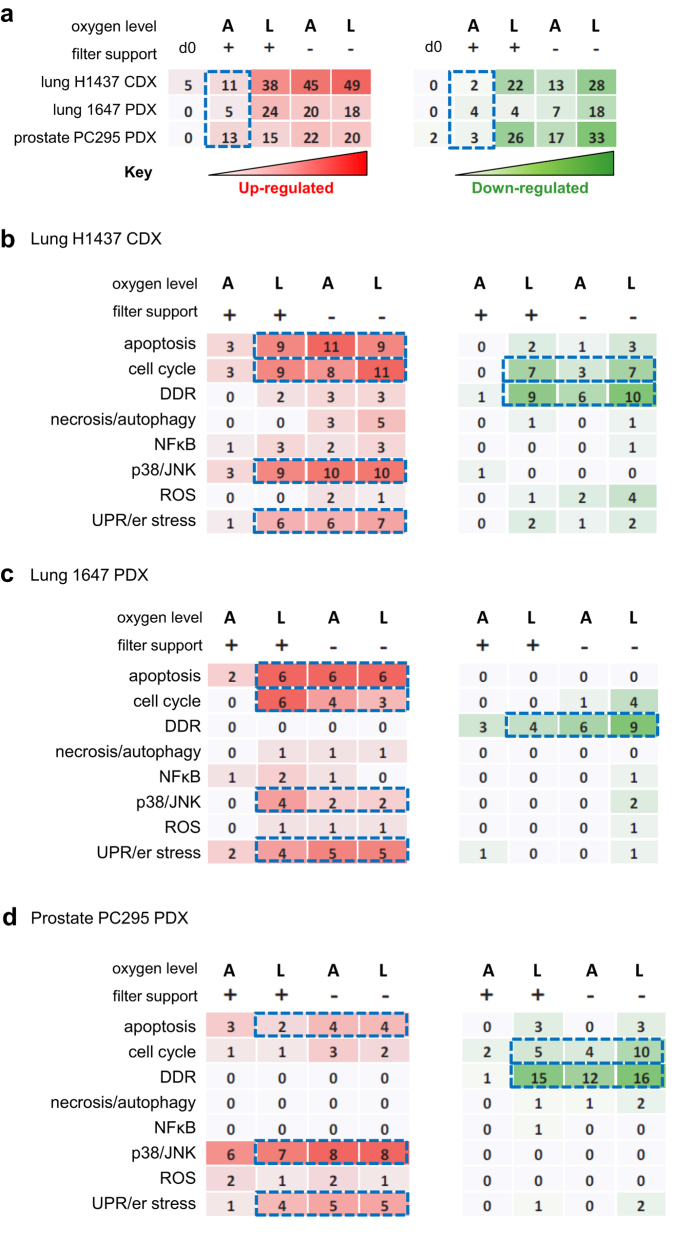
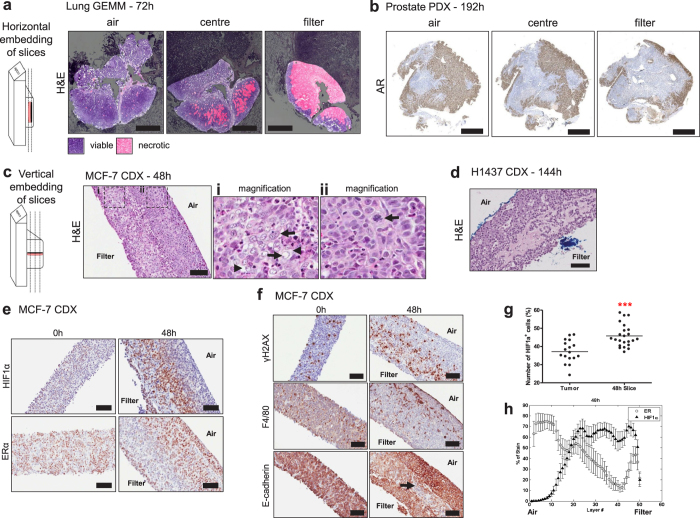
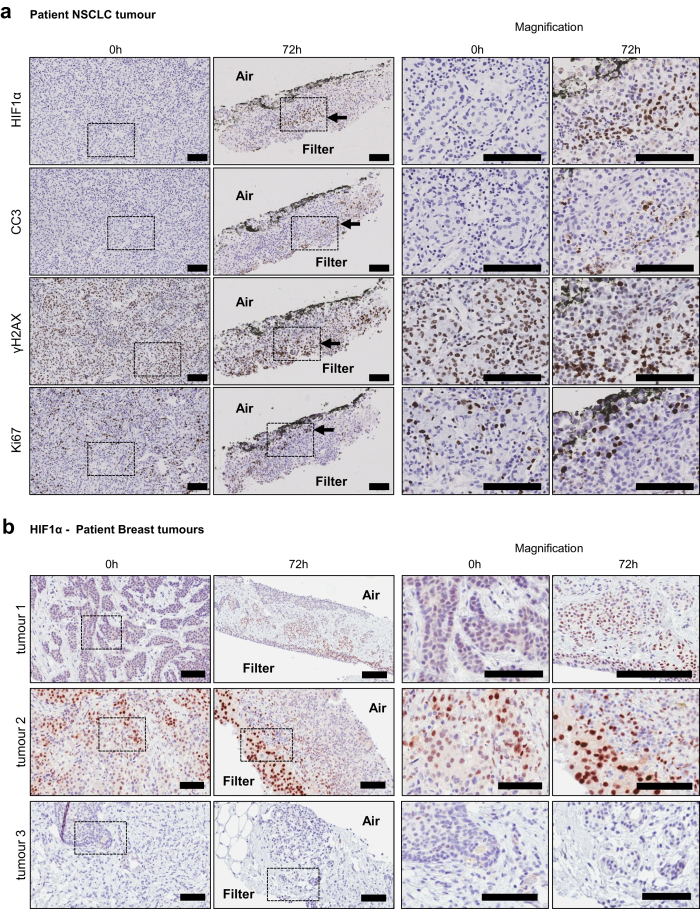
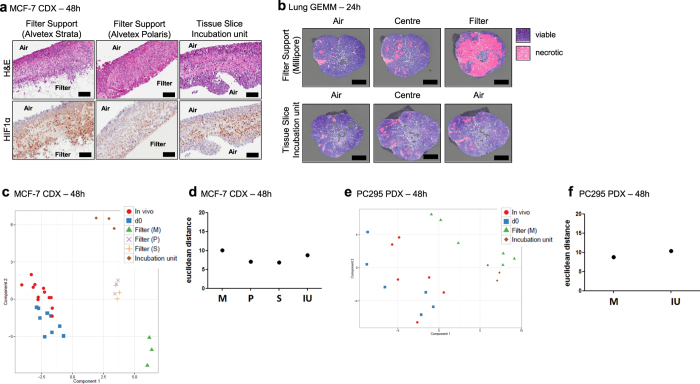
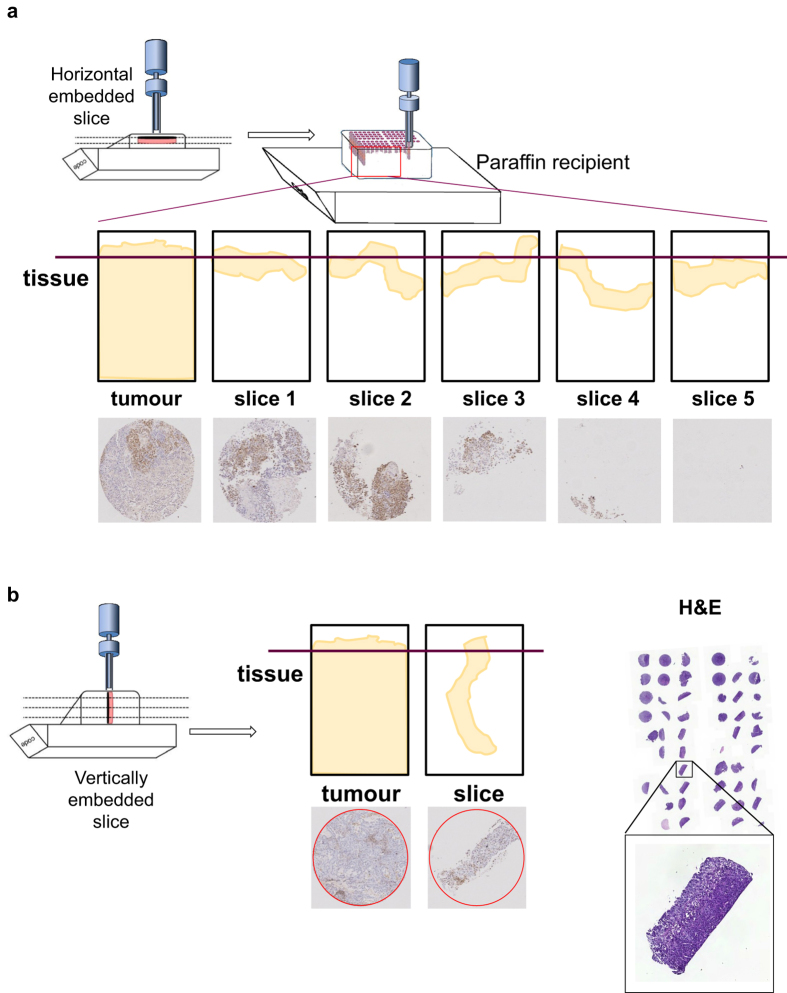
Precision-cut slices of in vivo tumours permit interrogation in vitro of heterogeneous cells from solid tumours together with their native microenvironment. They offer a low throughput but high content in vitro experimental platform. Using mouse models as surrogates for three common human solid tumours, we describe a standardised workflow for systematic comparison of tumour slice cultivation methods and a tissue microarray-based method to archive them. Cultivated slices were compared to their in vivo source tissue using immunohistochemical and transcriptional biomarkers, particularly of cellular stress. Mechanical slicing induced minimal stress. Cultivation of tumour slices required organotypic support materials and atmospheric oxygen for maintenance of integrity and was associated with significant temporal and loco-regional changes in protein expression, for example HIF-1α. We recommend adherence to the robust workflow described, with recognition of temporal-spatial changes in protein expression before interrogation of tumour slices by pharmacological or other means.
Figures
References
-
- McMillin D. W., Negri J. M. & Mitsiades C. S. The role of tumour-stromal interactions in modifying drug response: challenges and opportunities. Nat Rev Drug Discov 12, 217–228, doi: nrd387010.1038/nrd3870 (2013). - PubMed
-
- Junttila M. R. & de Sauvage F. J. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 501, 346–354, doi: nature1262610.1038/nature12626 (2013). - PubMed
-
- Wilding J. L. & Bodmer W. F. Cancer cell lines for drug discovery and development. Cancer Res 74, 2377–2384, doi: 0008-5472.CAN-13-297110.1158/0008-5472.CAN-13-2971 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
