

Ferredoxin 1b (Fdx1b) Is the Essential Mitochondrial Redox Partner for Cortisol Biosynthesis in Zebrafish
- PMID: 26650568
- PMCID: PMC4769370
- DOI: 10.1210/en.2015-1480
Ferredoxin 1b (Fdx1b) Is the Essential Mitochondrial Redox Partner for Cortisol Biosynthesis in Zebrafish
Abstract
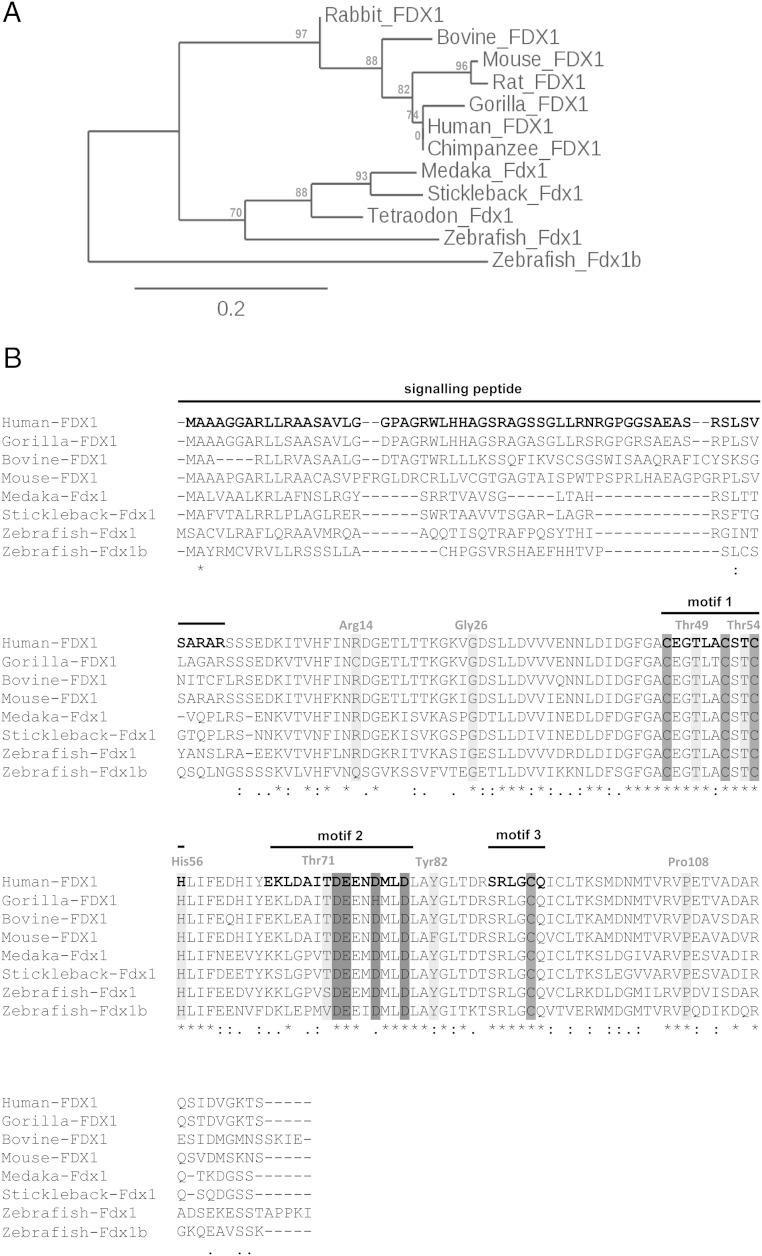
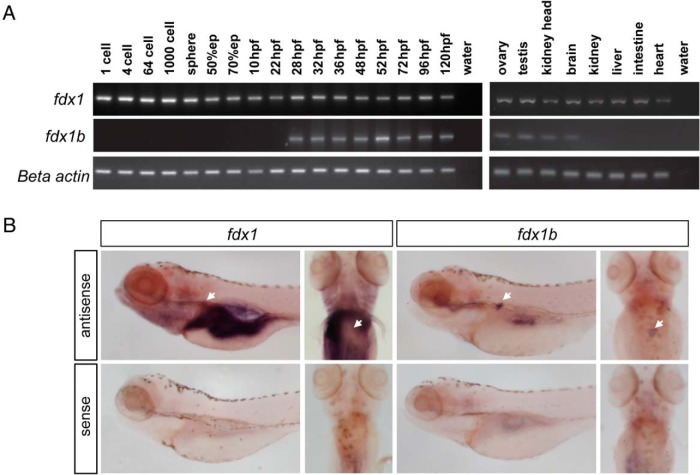
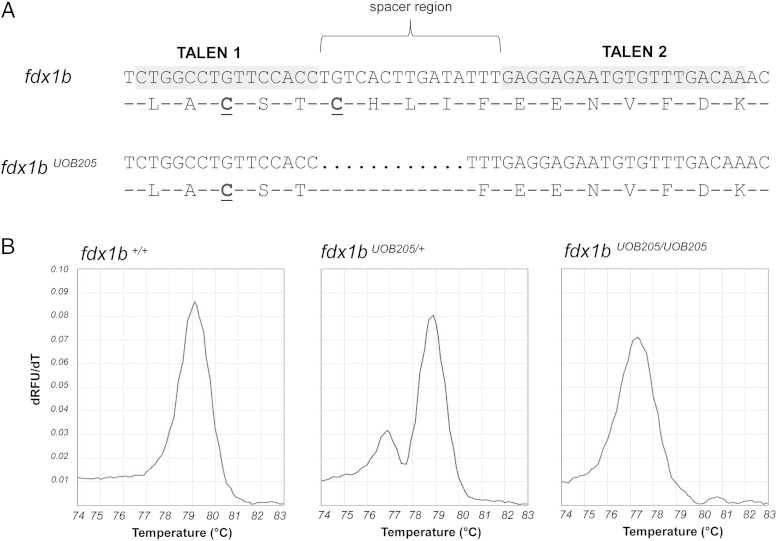
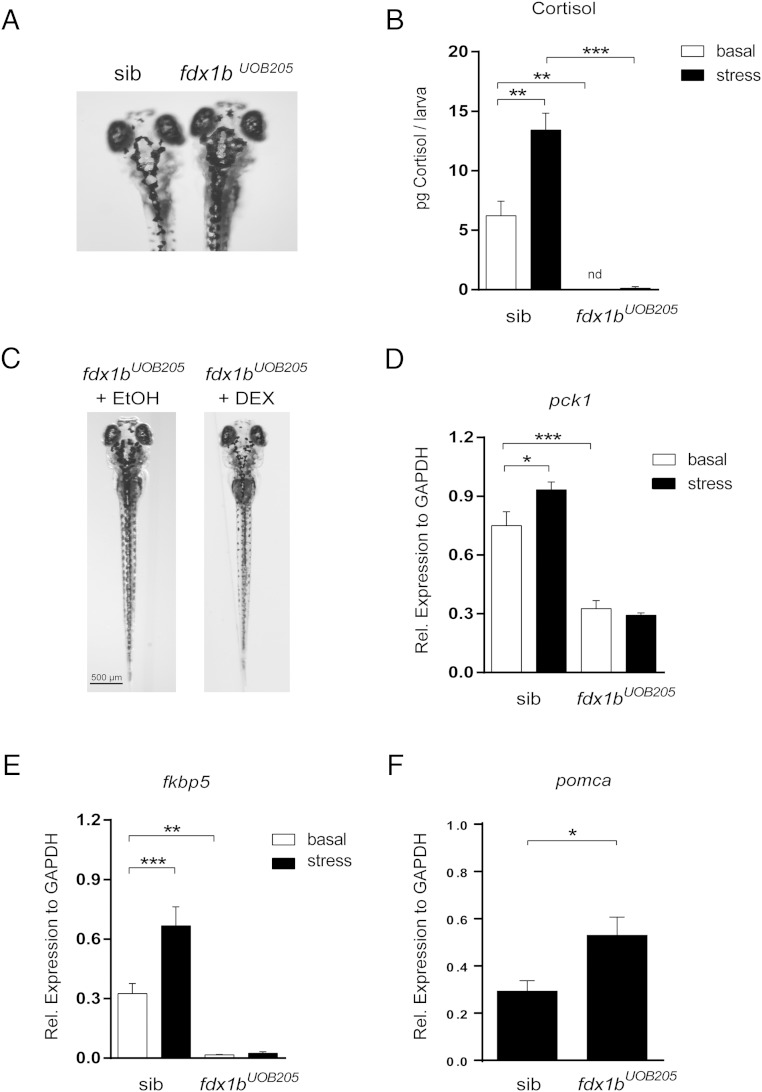
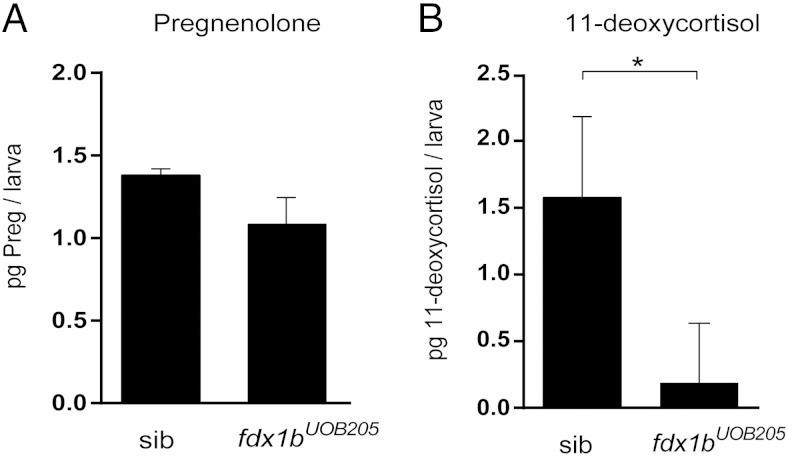
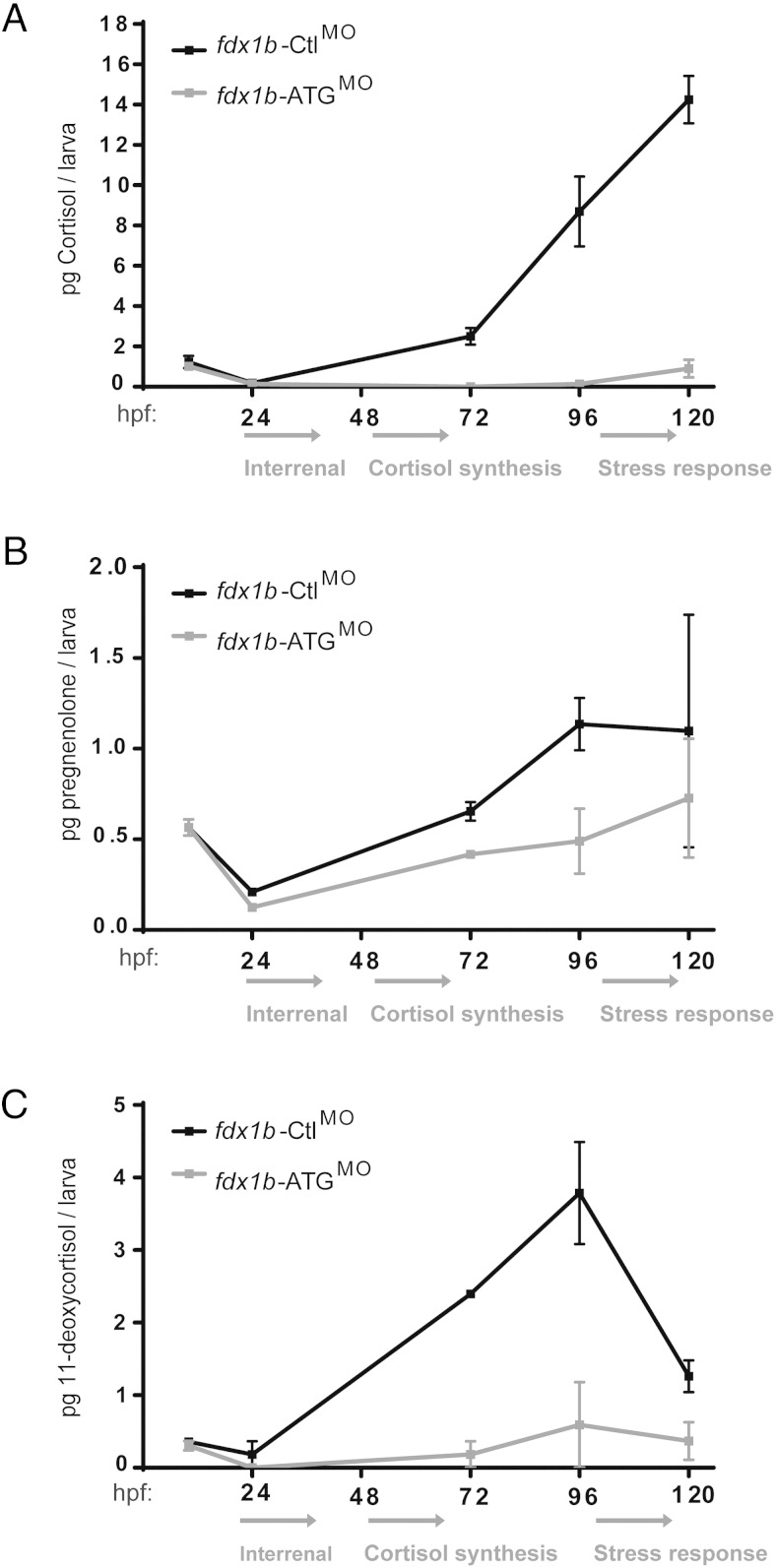
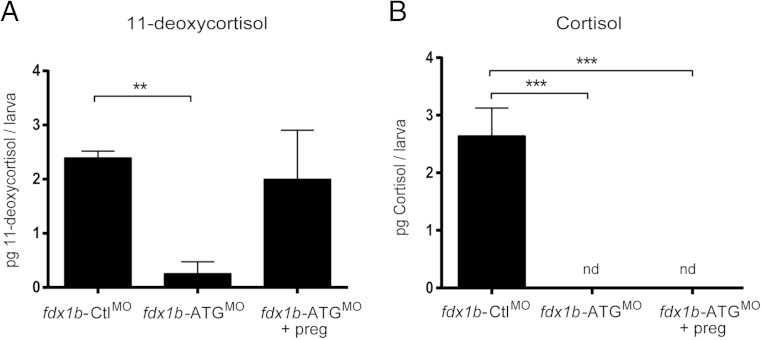
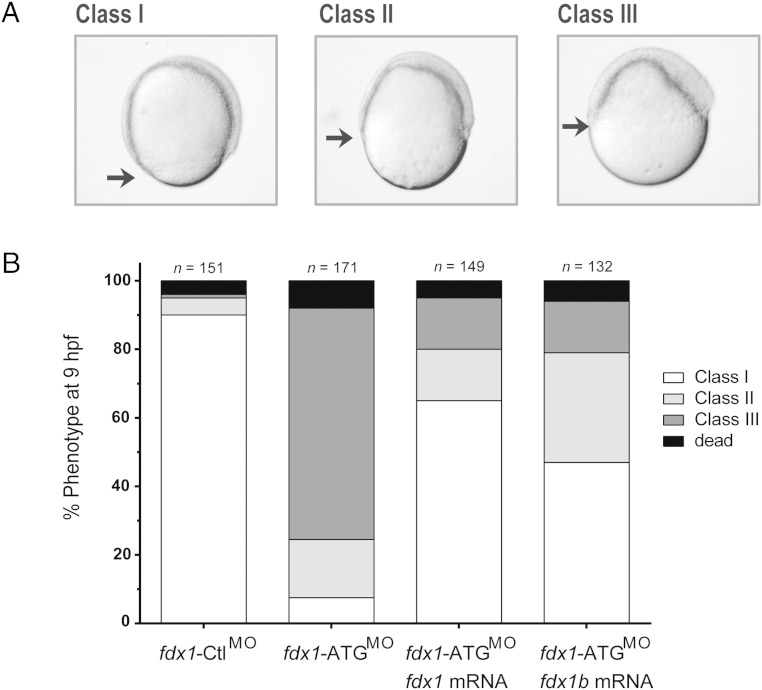
Mitochondrial cytochrome P450 (CYP) enzymes rely on electron transfer from the redox partner ferredoxin 1 (FDX1) for catalytic activity. Key steps in steroidogenesis require mitochondrial CYP enzymes and FDX1. Over 30 ferredoxin mutations have been explored in vitro; however, no spontaneously occurring mutations have been identified in humans leaving the impact of FDX1 on steroidogenesis in the whole organism largely unknown. Zebrafish are an important model to study human steroidogenesis, because they have similar steroid products and endocrine tissues. This study aimed to characterize the influence of ferredoxin on steroidogenic capacity in vivo by using zebrafish. Zebrafish have duplicate ferredoxin paralogs: fdx1 and fdx1b. Although fdx1 was observed throughout development and in most tissues, fdx1b was expressed after development of the zebrafish interrenal gland (counterpart to the mammalian adrenal gland). Additionally, fdx1b was restricted to adult steroidogenic tissues, such as the interrenal, gonads, and brain, suggesting that fdx1b was interacting with steroidogenic CYP enzymes. By using transcription activator-like effector nucleases, we generated fdx1b mutant zebrafish lines. Larvae with genetic disruption of fdx1b were morphologically inconspicuous. However, steroid hormone analysis by liquid chromatography tandem mass spectrometry revealed fdx1b mutants failed to synthesize glucocorticoids. Additionally, these mutants had an up-regulation of the hypothalamus-pituitary-interrenal axis and showed altered dark-light adaptation, suggesting impaired cortisol signaling. Antisense morpholino knockdown confirmed Fdx1b is required for de novo cortisol biosynthesis. In summary, by using zebrafish, we generated a ferredoxin knockout model system, which demonstrates for the first time the impact of mitochondrial redox regulation on glucocorticoid biosynthesis in vivo.
Figures
Comment in
-
The First Defect in Electron Transfer to Mitochondrial P450 Enzymes.Endocrinology. 2016 Mar;157(3):1003-6. doi: 10.1210/en.2016-1044. Endocrinology. 2016. PMID: 26919515 No abstract available.
References
-
- Miller WL. Minireview: regulation of steroidogenesis by electron transfer. Endocrinology. 2005;146:2544–2550. - PubMed
-
- Meyer J. Iron-sulfur protein folds, iron-sulfur chemistry, and evolution. J Biol Inorg Chem. 2008;13:157–170. - PubMed
-
- Ewen KM, Ringle M, Bernhardt R. Adrenodoxin–a versatile ferredoxin. IUBMB Life. 2012;64:506–512. - PubMed
-
- Miller WL. Steroid hormone synthesis in mitochondria. Mol Cell Endocrinol. 2013;379:62–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
