

Anxiety-Like Behavioural Inhibition Is Normative under Environmental Threat-Reward Correlations
- PMID: 26650585
- PMCID: PMC4674090
- DOI: 10.1371/journal.pcbi.1004646
Anxiety-Like Behavioural Inhibition Is Normative under Environmental Threat-Reward Correlations
Abstract
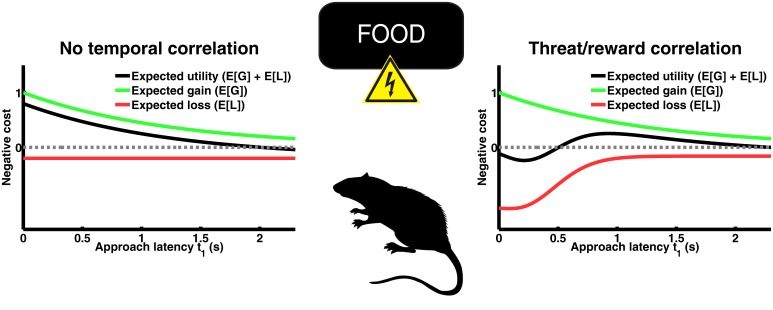
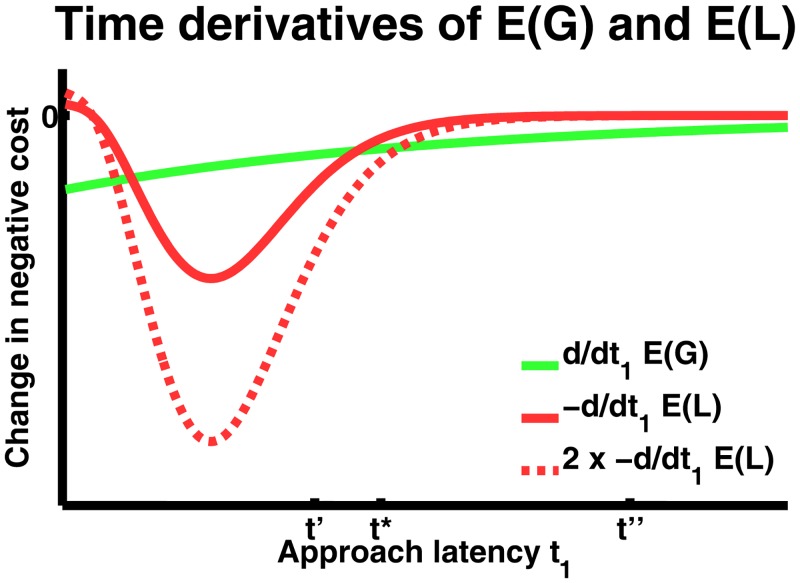
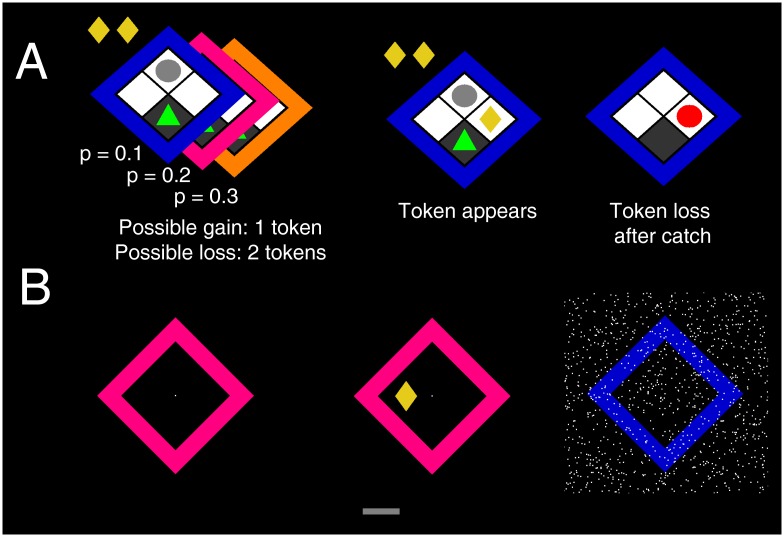
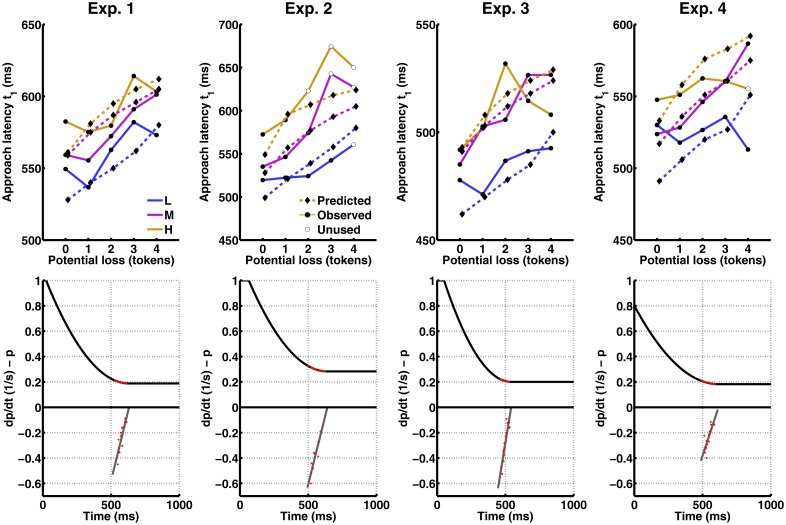
Behavioural inhibition is a key anxiety-like behaviour in rodents and humans, distinct from avoidance of danger, and reduced by anxiolytic drugs. In some situations, it is not clear how behavioural inhibition minimises harm or maximises benefit for the agent, and can even appear counterproductive. Extant explanations of this phenomenon make use of descriptive models but do not provide a formal assessment of its adaptive value. This hampers a better understanding of the neural computations underlying anxiety behaviour. Here, we analyse a standard rodent anxiety model, the operant conflict test. We harvest Bayesian Decision Theory to show that behavioural inhibition normatively arises as cost-minimising strategy in temporally correlated environments. Importantly, only if behavioural inhibition is aimed at minimising cost, it depends on probability and magnitude of threat. Harnessing a virtual computer game, we test model predictions in four experiments with human participants. Humans exhibit behavioural inhibition with a strong linear dependence on threat probability and magnitude. Strikingly, inhibition occurs before motor execution and depends on the virtual environment, thus likely resulting from a neural optimisation process rather than a pre-programmed mechanism. Individual trait anxiety scores predict behavioural inhibition, underlining the validity of this anxiety model. These findings put anxiety behaviour into the context of cost-minimisation and optimal inference, and may ultimately pave the way towards a mechanistic understanding of the neural computations gone awry in human anxiety disorder.
Conflict of interest statement
The author has declared that no competing interests exist.
Figures

References
-
- Gray JA, McNaughton N. The neuropsychology of anxiety: an enquiry into the functions of the septo-hippocampal system. vol. no. 33 2nd ed Oxford: Oxford University Press; 2000. Available from: http://www.loc.gov/catdir/enhancements/fy0639/2001280232-d.html.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
