

Non-genetic Purification of Ventricular Cardiomyocytes from Differentiating Embryonic Stem Cells through Molecular Beacons Targeting IRX-4
- PMID: 26651608
- PMCID: PMC4682289
- DOI: 10.1016/j.stemcr.2015.10.021
Non-genetic Purification of Ventricular Cardiomyocytes from Differentiating Embryonic Stem Cells through Molecular Beacons Targeting IRX-4
Abstract
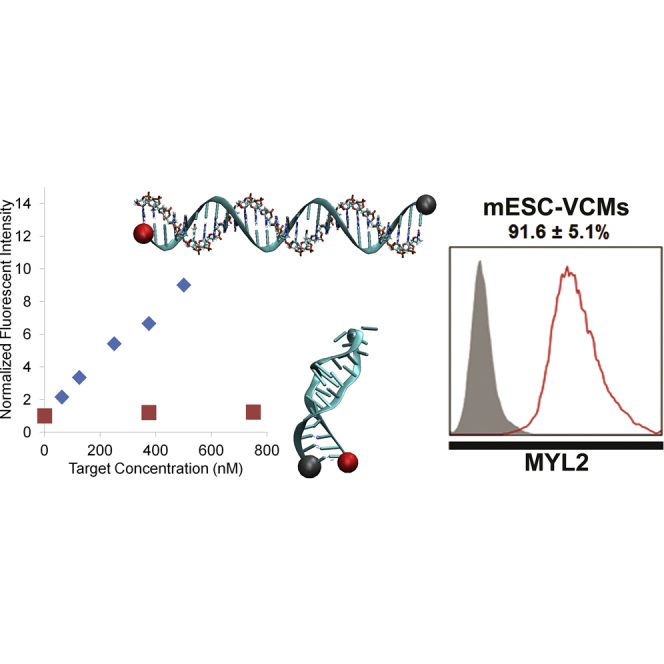
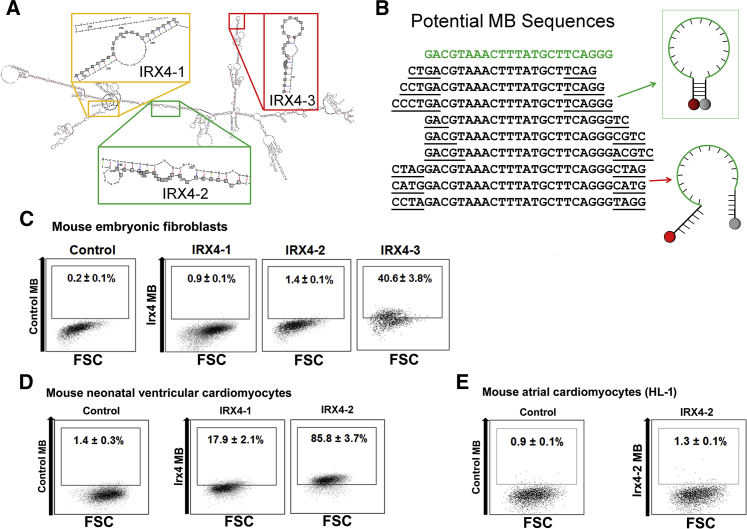
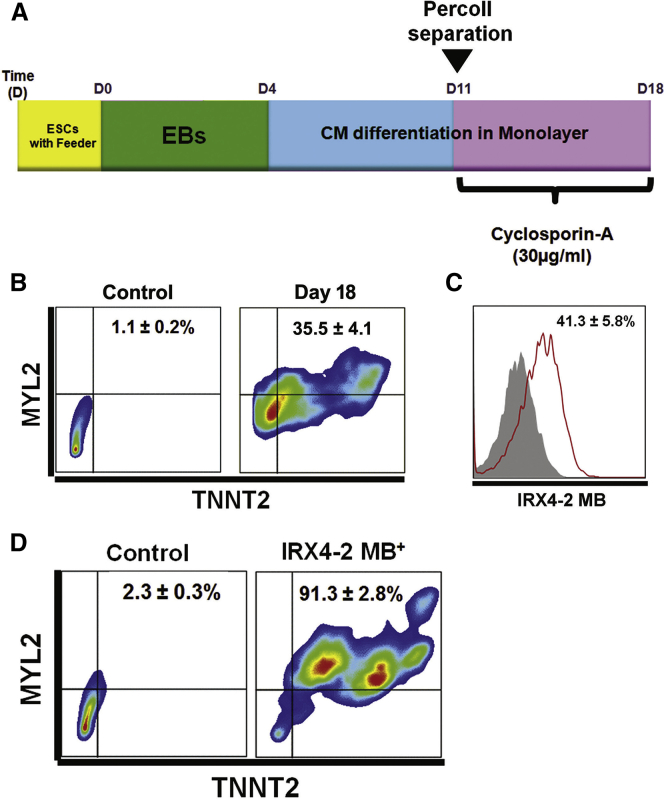
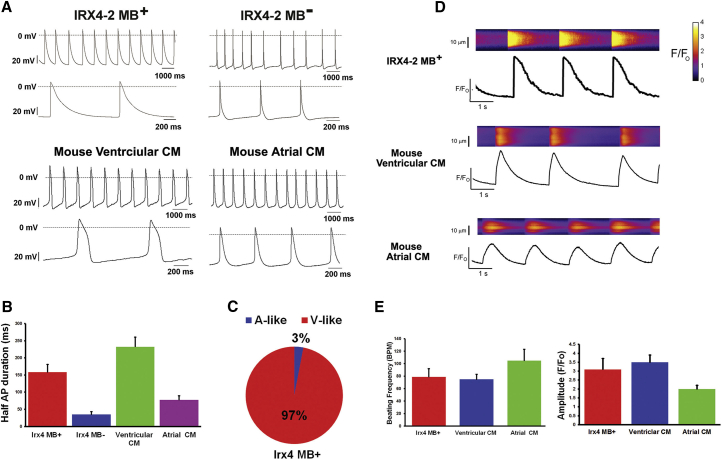
Isolation of ventricular cardiomyocytes (vCMs) has been challenging due to the lack of specific surface markers. Here we show that vCMs can be purified from differentiating mouse embryonic stem cells (mESCs) using molecular beacons (MBs) targeting specific intracellular mRNAs. We designed MBs (IRX4 MBs) to target mRNA encoding Iroquois homeobox protein 4 (Irx4), a transcription factor specific for vCMs. To purify mESC vCMs, IRX4 MBs were delivered into cardiomyogenically differentiating mESCs, and IRX4 MBs-positive cells were FACS-sorted. We found that, of the cells isolated, ~98% displayed vCM-like action potentials by electrophysiological analyses. These MB-purified vCMs continuously maintained their CM characteristics as verified by spontaneous beating, Ca(2+) transient, and expression of vCM-specific proteins. Our study shows the feasibility of isolating pure vCMs via cell sorting without modifying host genes. The homogeneous and functional ventricular CMs generated via the MB-based method can be useful for disease investigation, drug discovery, and cell-based therapies.
Keywords: IRX4; cell selection; molecular beacons; pluripotent stem cell; ventricular cardiomyocyte.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Purification of cardiomyocytes from differentiating pluripotent stem cells using molecular beacons that target cardiomyocyte-specific mRNA.Circulation. 2013 Oct 22;128(17):1897-909. doi: 10.1161/CIRCULATIONAHA.113.004228. Epub 2013 Aug 30. Circulation. 2013. PMID: 23995537 Free PMC article.
-
Derivation of functional ventricular cardiomyocytes using endogenous promoter sequence from murine embryonic stem cells.Stem Cell Res. 2012 Jan;8(1):49-57. doi: 10.1016/j.scr.2011.08.004. Epub 2011 Aug 22. Stem Cell Res. 2012. PMID: 22099020 Free PMC article.
-
Characterization of Ventricular and Atrial Cardiomyocyte Subtypes from Human-Induced Pluripotent Stem Cells.Methods Mol Biol. 2021;2320:135-149. doi: 10.1007/978-1-0716-1484-6_14. Methods Mol Biol. 2021. PMID: 34302655
-
Molecular beacon-based detection and isolation of working-type cardiomyocytes derived from human pluripotent stem cells.Biomaterials. 2015 May;50:176-85. doi: 10.1016/j.biomaterials.2015.01.043. Epub 2015 Feb 19. Biomaterials. 2015. PMID: 25736507 Free PMC article.
-
Generation and purification of human stem cell-derived cardiomyocytes.Differentiation. 2016 Apr-Jun;91(4-5):126-38. doi: 10.1016/j.diff.2016.01.001. Epub 2016 Feb 22. Differentiation. 2016. PMID: 26915912 Review.
Cited by
-
KCNQ1-deficient and KCNQ1-mutant human embryonic stem cell-derived cardiomyocytes for modeling QT prolongation.Stem Cell Res Ther. 2022 Jun 28;13(1):287. doi: 10.1186/s13287-022-02964-3. Stem Cell Res Ther. 2022. PMID: 35765105 Free PMC article.
-
Probing early heart development to instruct stem cell differentiation strategies.Dev Dyn. 2016 Dec;245(12):1130-1144. doi: 10.1002/dvdy.24441. Epub 2016 Oct 3. Dev Dyn. 2016. PMID: 27580352 Free PMC article. Review.
-
Cardiomyocyte precursors generated by direct reprogramming and molecular beacon selection attenuate ventricular remodeling after experimental myocardial infarction.Stem Cell Res Ther. 2023 Oct 15;14(1):296. doi: 10.1186/s13287-023-03519-w. Stem Cell Res Ther. 2023. PMID: 37840130 Free PMC article.
-
Iroquois Homeodomain transcription factors in ventricular conduction system and arrhythmia.Int J Med Sci. 2018 May 22;15(8):808-815. doi: 10.7150/ijms.25140. eCollection 2018. Int J Med Sci. 2018. PMID: 30008591 Free PMC article. Review.
-
Technical approaches to induce selective cell death of pluripotent stem cells.Cell Mol Life Sci. 2017 Jul;74(14):2601-2611. doi: 10.1007/s00018-017-2486-0. Epub 2017 Feb 28. Cell Mol Life Sci. 2017. PMID: 28246701 Free PMC article. Review.
References
-
- Bao Z.-Z., Bruneau B.G., Seidman J.G., Seidman C.E., Cepko C.L. Regulation of chamber-specific gene expression in the developing heart by Irx4. Science. 1999;283:1161–1164. - PubMed
-
- Brundel B.J.J.M., Henning R.H., Ke L., van Gelder I.C., Crijns H.J.G.M., Kampinga H.H. Heat shock protein upregulation protects against pacing-induced myolysis in HL-1 atrial myocytes and in human atrial fibrillation. J. Mol. Cell. Cardiol. 2006;41:555–562. - PubMed
-
- Bruneau B.G., Bao Z.-Z., Tanaka M., Schott J.-J., Izumo S., Cepko C.L., Seidman J.G., Seidman C.E. Cardiac expression of the ventricle-specific homeobox gene Irx4 is modulated by Nkx2-5 and dHand. Dev. Biol. 2000;217:266–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
