

A Single Bacterial Immune Evasion Strategy Dismantles Both MyD88 and TRIF Signaling Pathways Downstream of TLR4
- PMID: 26651944
- PMCID: PMC4685476
- DOI: 10.1016/j.chom.2015.11.006
A Single Bacterial Immune Evasion Strategy Dismantles Both MyD88 and TRIF Signaling Pathways Downstream of TLR4
Abstract
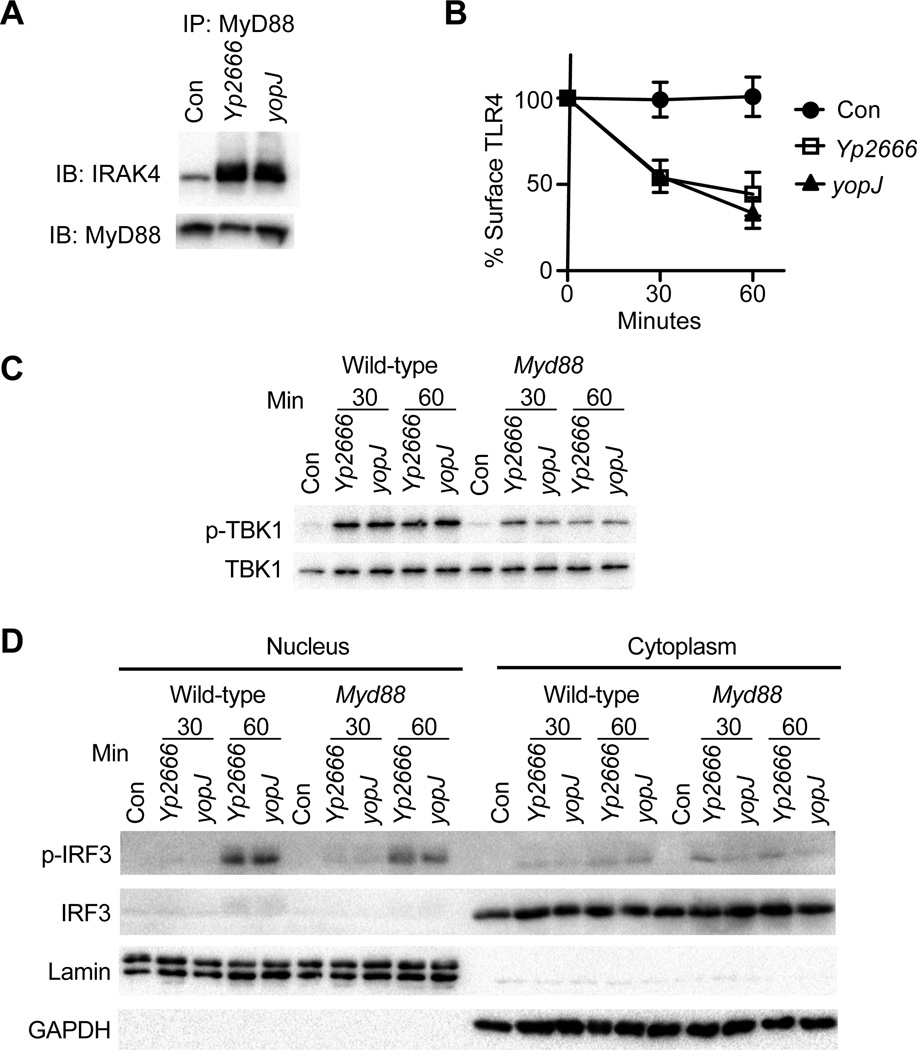
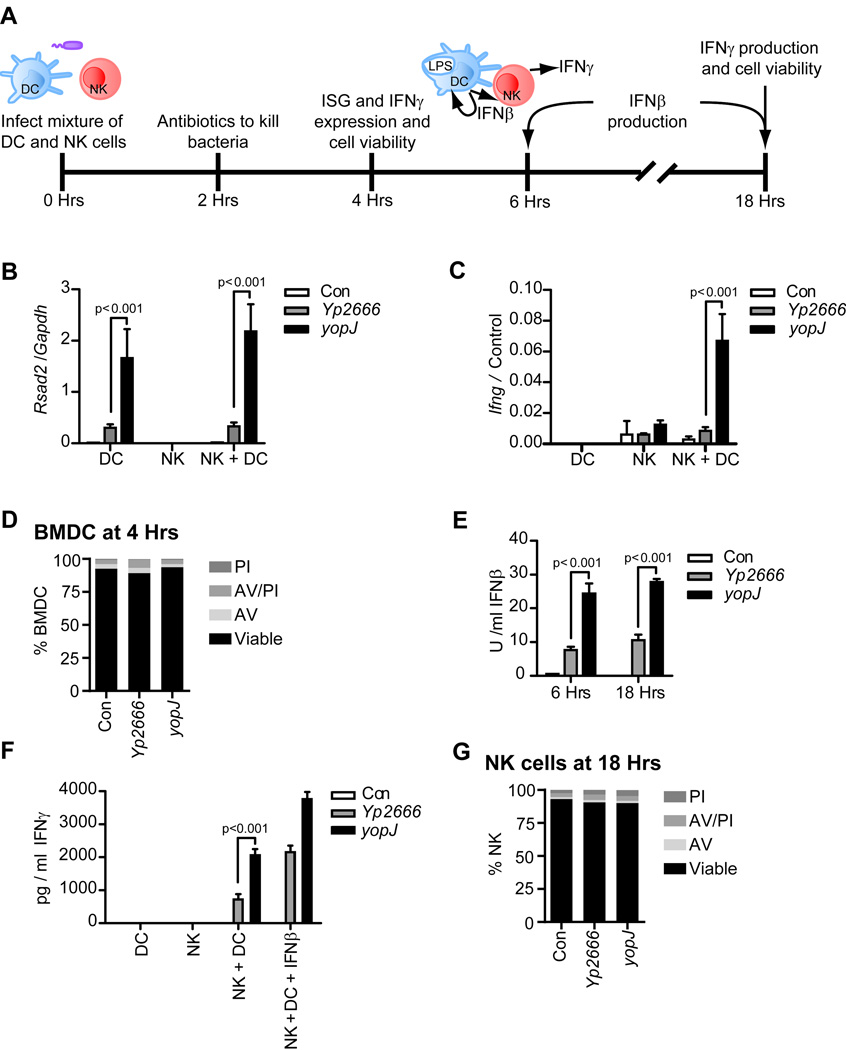
During bacterial infections, Toll-like receptor 4 (TLR4) signals through the MyD88- and TRIF-dependent pathways to promote pro-inflammatory and interferon (IFN) responses, respectively. Bacteria can inhibit the MyD88 pathway, but if the TRIF pathway is also targeted is unclear. We demonstrate that, in addition to MyD88, Yersinia pseudotuberculosis inhibits TRIF signaling through the type III secretion system effector YopJ. Suppression of TRIF signaling occurs during dendritic cell (DC) and macrophage infection and prevents expression of type I IFN and pro-inflammatory cytokines. YopJ-mediated inhibition of TRIF prevents DCs from inducing natural killer (NK) cell production of antibacterial IFNγ. During infection of DCs, YopJ potently inhibits MAPK pathways but does not prevent activation of IKK- or TBK1-dependent pathways. This singular YopJ activity efficiently inhibits TLR4 transcription-inducing activities, thus illustrating a simple means by which pathogens impede innate immunity.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures




References
-
- Baxt LA, Garza-Mayers AC, Goldberg MB. Bacterial subversion of host innate immune pathways. Science. 2013;340:697–701. - PubMed
MeSH terms
Substances
Grants and funding
- R01 AI093589/AI/NIAID NIH HHS/United States
- R56 AI093589/AI/NIAID NIH HHS/United States
- R56 AI103082/AI/NIAID NIH HHS/United States
- R01 AI103062/AI/NIAID NIH HHS/United States
- AI113141/AI/NIAID NIH HHS/United States
- DK102317-01/DK/NIDDK NIH HHS/United States
- P30 DK034854/DK/NIDDK NIH HHS/United States
- R01 AI113166/AI/NIAID NIH HHS/United States
- K99 AI072955/AI/NIAID NIH HHS/United States
- P30 DK34854/DK/NIDDK NIH HHS/United States
- AI113166/AI/NIAID NIH HHS/United States
- R00 AI072955/AI/NIAID NIH HHS/United States
- F32 DK102317/DK/NIDDK NIH HHS/United States
- R56 AI113141/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
