

SIAH ubiquitin ligases regulate breast cancer cell migration and invasion independent of the oxygen status
- PMID: 26654769
- PMCID: PMC4825722
- DOI: 10.1080/15384101.2015.1104441
SIAH ubiquitin ligases regulate breast cancer cell migration and invasion independent of the oxygen status
Abstract
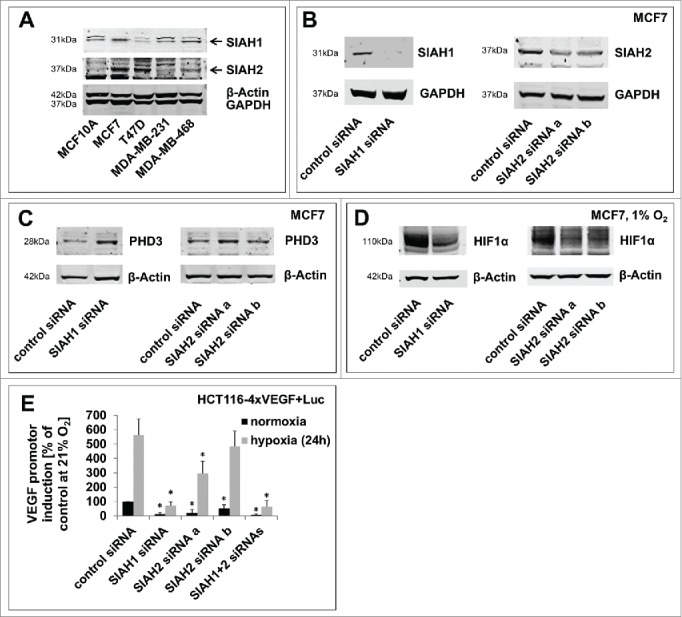
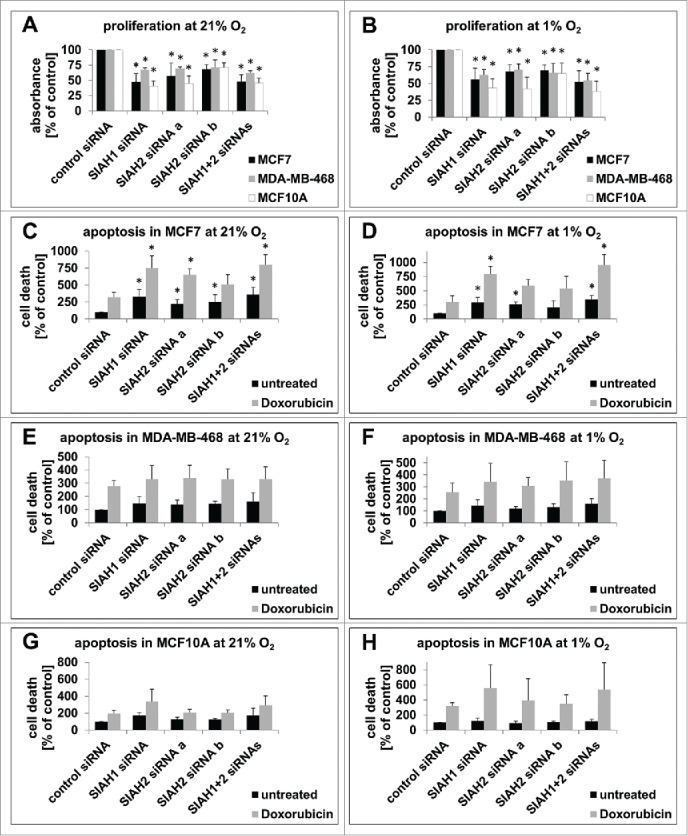
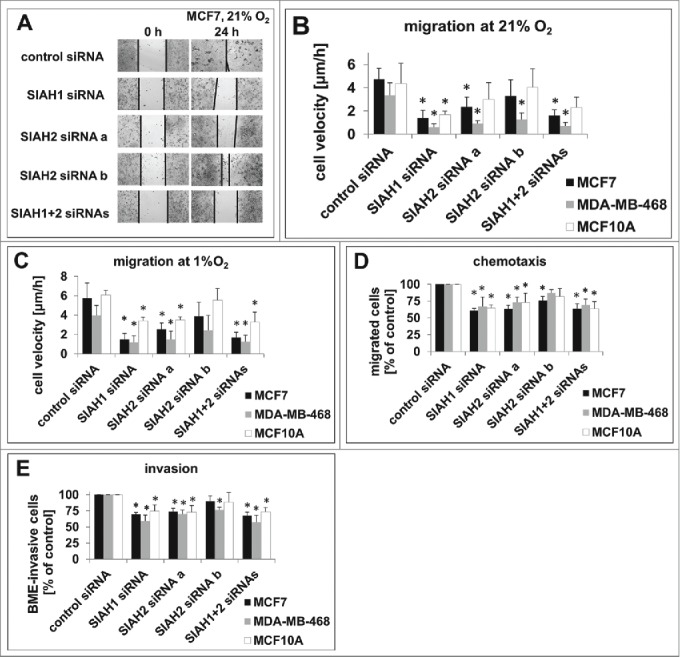
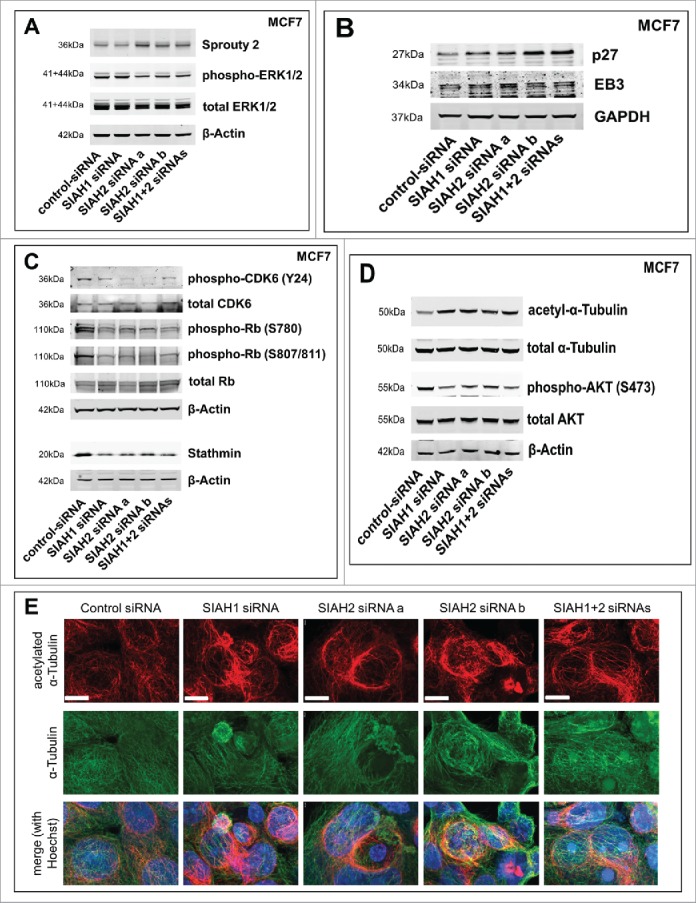
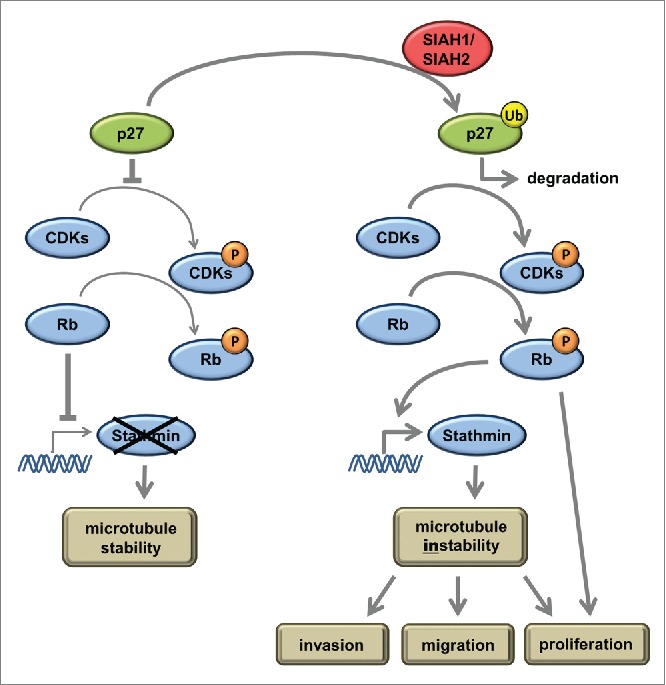
Seven-in-absentia homolog (SIAH) proteins are evolutionary conserved RING type E3 ubiquitin ligases responsible for the degradation of key molecules regulating DNA damage response, hypoxic adaptation, apoptosis, angiogenesis, and cell proliferation. Many studies suggest a tumorigenic role for SIAH2. In breast cancer patients SIAH2 expression levels correlate with cancer aggressiveness and overall patient survival. In addition, SIAH inhibition reduced metastasis in melanoma. The role of SIAH1 in breast cancer is still ambiguous; both tumorigenic and tumor suppressive functions have been reported. Other studies categorized SIAH ligases as either pro- or antimigratory, while the significance for metastasis is largely unknown. Here, we re-evaluated the effects of SIAH1 and SIAH2 depletion in breast cancer cell lines, focusing on migration and invasion. We successfully knocked down SIAH1 and SIAH2 in several breast cancer cell lines. In luminal type MCF7 cells, this led to stabilization of the SIAH substrate Prolyl Hydroxylase Domain protein 3 (PHD3) and reduced Hypoxia-Inducible Factor 1α (HIF1α) protein levels. Both the knockdown of SIAH1 or SIAH2 led to increased apoptosis and reduced proliferation, with comparable effects. These results point to a tumor promoting role for SIAH1 in breast cancer similar to SIAH2. In addition, depletion of SIAH1 or SIAH2 also led to decreased cell migration and invasion in breast cancer cells. SIAH knockdown also controlled microtubule dynamics by markedly decreasing the protein levels of stathmin, most likely via p27(Kip1). Collectively, these results suggest that both SIAH ligases promote a migratory cancer cell phenotype and could contribute to metastasis in breast cancer.
Keywords: SIAH ubiquitin ligases; SIAH1; SIAH2; breast cancer; invasion; metastasis; migration; p27Kip1; seven-in-absentia; stathmin.
Figures
References
-
- Hu G, Zhang S, Vidal M, Baer JL, Xu T, Fearon ER. Mammalian homologs of seven in absentia regulate DCC via the ubiquitin-proteasome pathway. Genes & Dev 1997; 11:2701-14; PMID:9334332; http://dx.doi.org/ 10.1101/gad.11.20.2701 - DOI - PMC - PubMed
-
- Roperch JP, Lethrone F, Prieur S, Piouffre L, Israeli D, Tuynder M, Nemani M, Pasturaud P, Gendron MC, Dausset J, et al.. SIAH-1 promotes apoptosis and tumor suppression through a network involving the regulation of protein folding, unfolding, and trafficking: identification of common effectors with p53 and p21(Waf1). Proc Natl Acad Sci U S A 1999; 96:8070-3; PMID:10393949; http://dx.doi.org/ 10.1073/pnas.96.14.8070 - DOI - PMC - PubMed
-
- Ahmed AU, Schmidt RL, Park CH, Reed NR, Hesse SE, Thomas CF, Molina JR, Deschamps C, Yang P, Aubry MC, et al.. Effect of disrupting seven-in-absentia homolog 2 function on lung cancer cell growth. J Natl Cancer Inst 2008; 100:1606-29; PMID:19001609; http://dx.doi.org/ 10.1093/jnci/djn365 - DOI - PMC - PubMed
-
- Behling KC, Tang A, Freydin B, Chervoneva I, Kadakia S, Schwartz GF, Rui H, Witkiewicz AK. Increased SIAH expression predicts ductal carcinoma in situ (DCIS) progression to invasive carcinoma. Breast Cancer Res Treat 2011; 129:717-24; PMID:21088888; http://dx.doi.org/ 10.1007/s10549-010-1254-8 - DOI - PMC - PubMed
-
- Qi J, Nakayama K, Cardiff RD, Borowsky AD, Kaul K, Williams R, Krajewski S, Mercola D, Carpenter PM, Bowtell D, et al.. Siah2-dependent concerted activity of HIF and FoxA2 regulates formation of neuroendocrine phenotype and neuroendocrine prostate tumors. Cancer Cell 2010; 18:23-38; PMID:20609350; http://dx.doi.org/ 10.1016/j.ccr.2010.05.024 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous