

The centrosome is an actin-organizing centre
- PMID: 26655833
- PMCID: PMC4880044
- DOI: 10.1038/ncb3285
The centrosome is an actin-organizing centre
Abstract
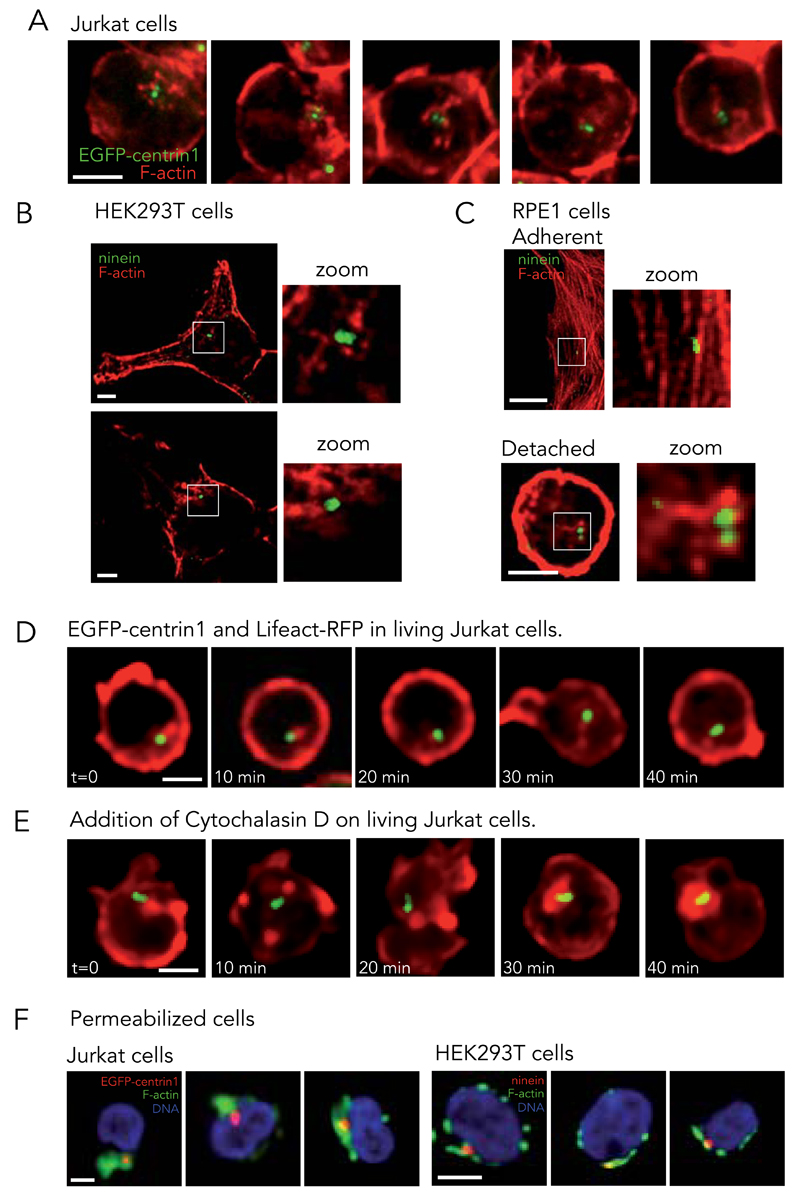
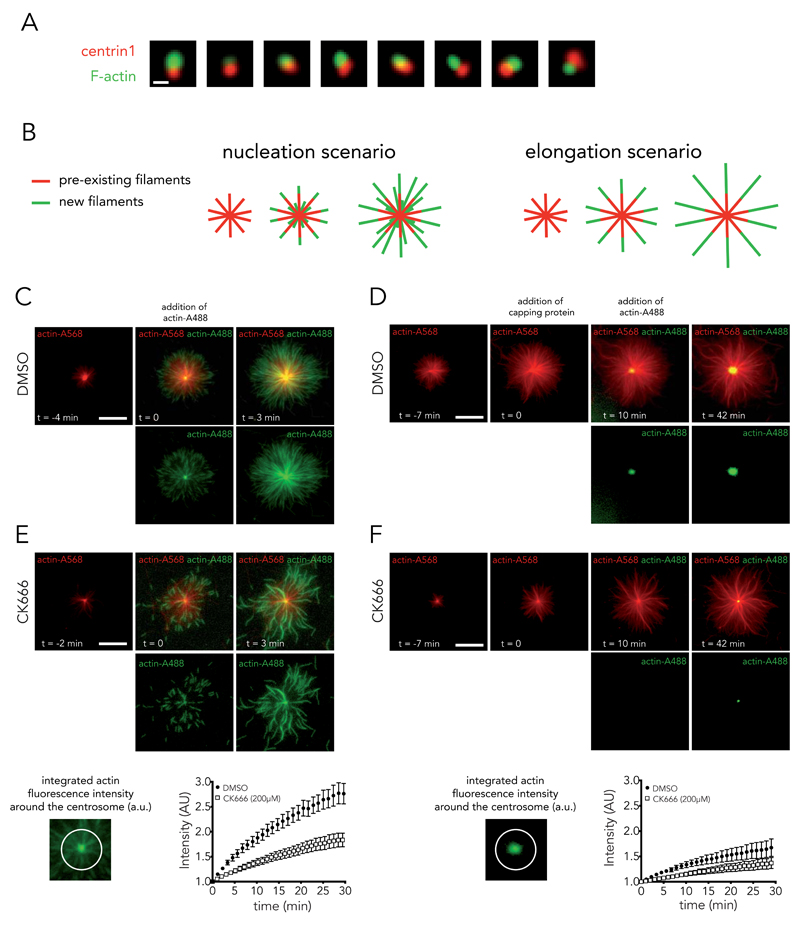
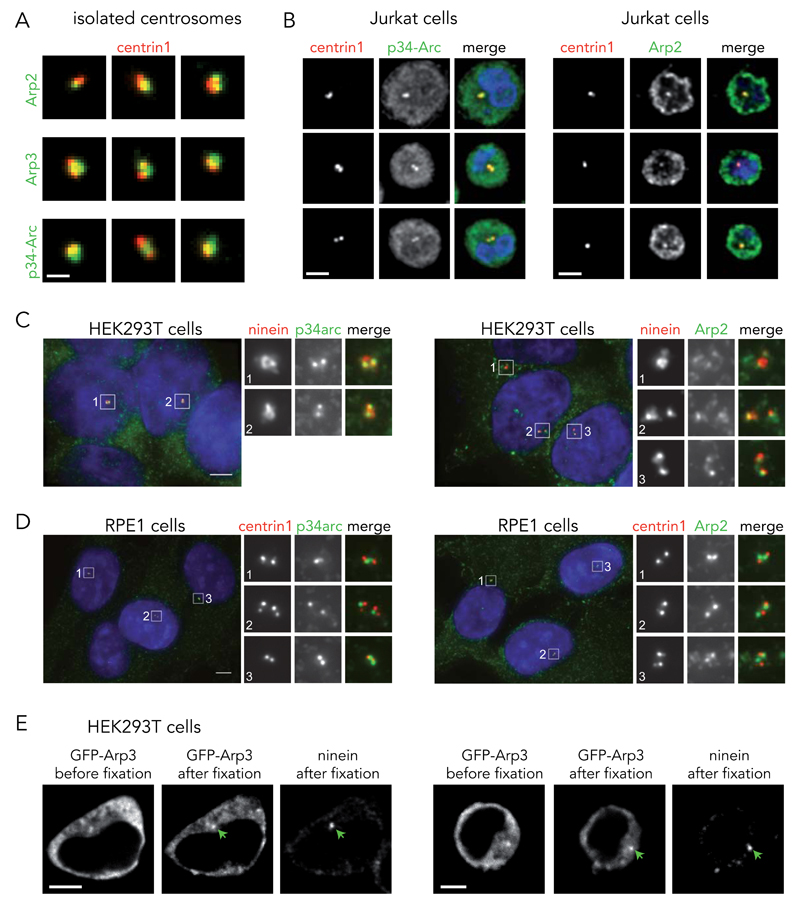
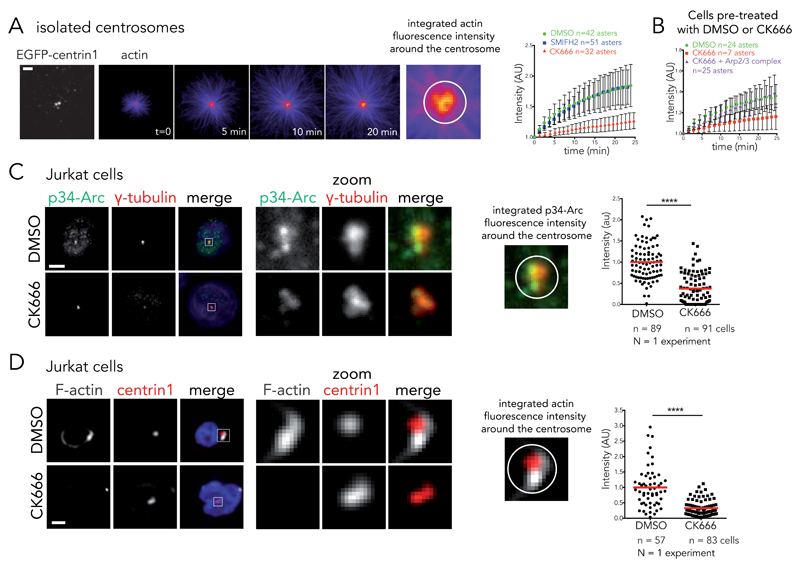
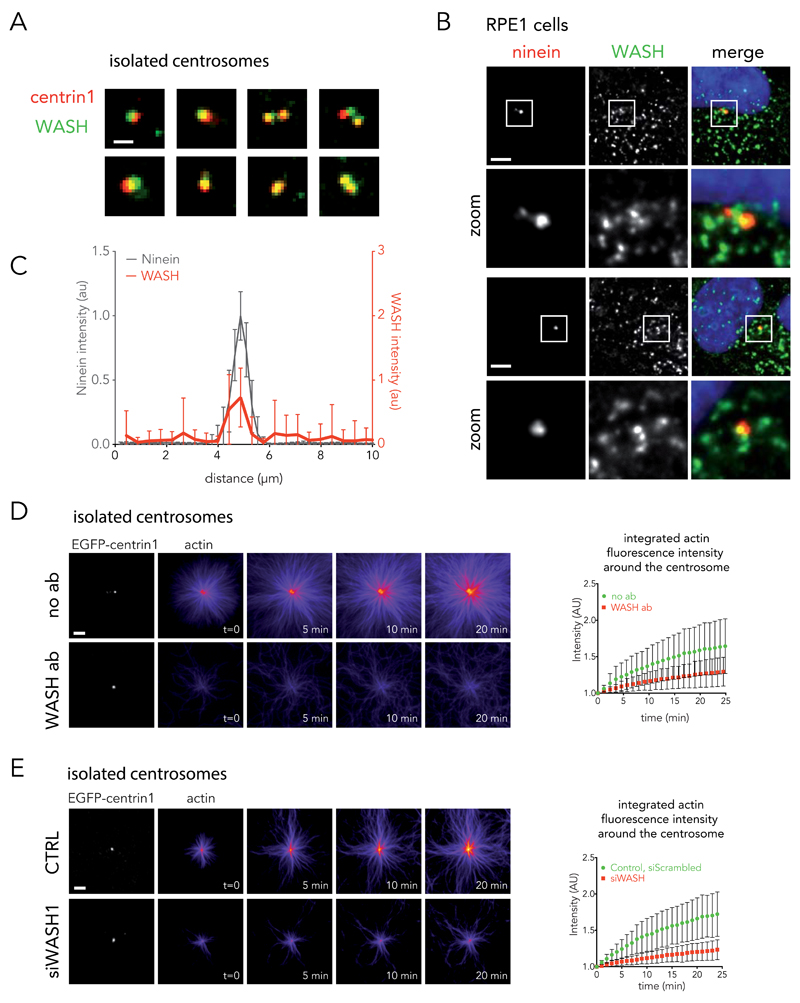
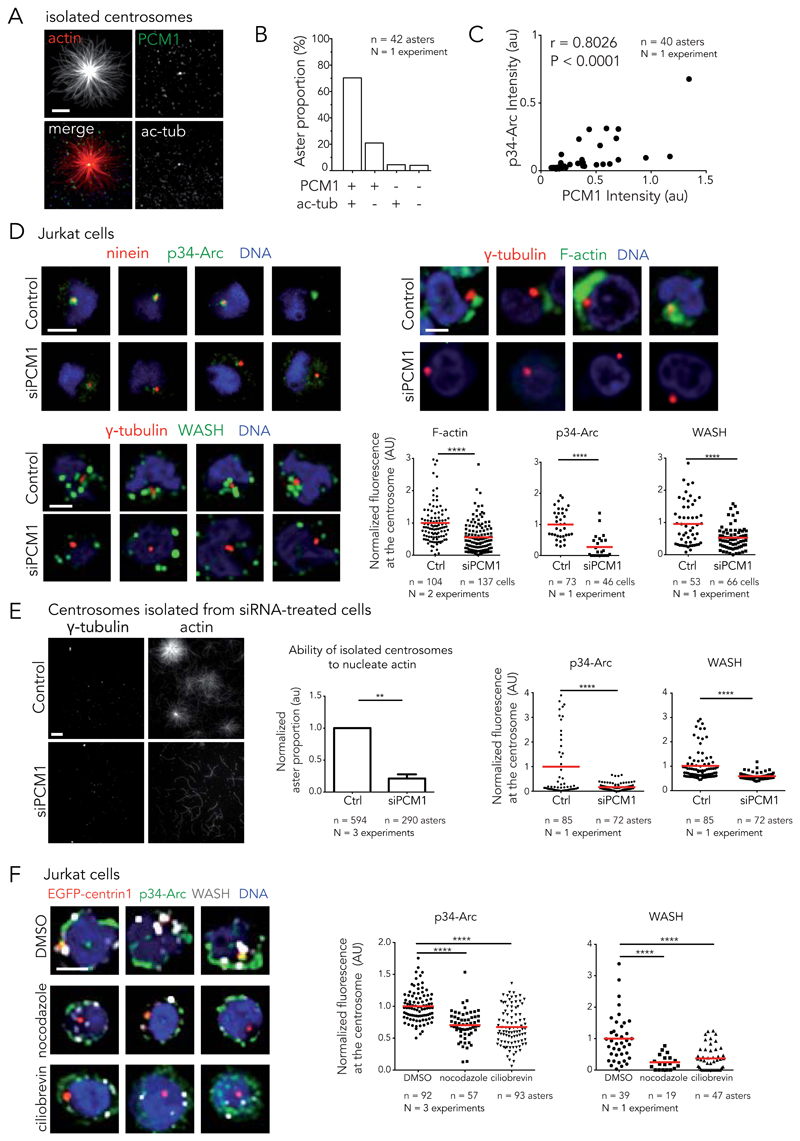
Microtubules and actin filaments are the two main cytoskeleton networks supporting intracellular architecture and cell polarity. The centrosome nucleates and anchors microtubules and is therefore considered to be the main microtubule-organizing centre. However, recurring, yet unexplained, observations have pointed towards a connection between the centrosome and actin filaments. Here we have used isolated centrosomes to demonstrate that the centrosome can directly promote actin-filament assembly. A cloud of centrosome-associated actin filaments could be identified in living cells as well. Actin-filament nucleation at the centrosome was mediated by the nucleation-promoting factor WASH in combination with the Arp2/3 complex. Pericentriolar material 1 (PCM1) seemed to modulate the centrosomal actin network by regulating Arp2/3 complex and WASH recruitment to the centrosome. Hence, our results reveal an additional facet of the centrosome as an intracellular organizer and provide mechanistic insights into how the centrosome can function as an actin-filament-organizing centre.
Conflict of interest statement
Competing financial interests
The authors declare no competing financial interests.
Figures

Comment in
-
Cytoskeleton: A new face of the centrosome.Nat Rev Mol Cell Biol. 2016 Feb;17(2):66-7. doi: 10.1038/nrm.2015.26. Epub 2015 Dec 23. Nat Rev Mol Cell Biol. 2016. PMID: 26695193 No abstract available.
References
-
- Bornens M. Organelle positioning and cell polarity. Nat Rev Mol Cell Biol. 2008;9:874–886. - PubMed
-
- Etienne-Manneville S. Actin and microtubules in cell motility: which one is in control? Traffic. 2004;5:470–7. - PubMed
-
- Rodriguez OC, et al. Conserved microtubule – actin interactions in cell movement and morphogenesis. Nat Cell Biol. 2003;5:599–609. - PubMed
-
- Coles CH, Bradke F. Coordinating Neuronal Actin–Microtubule Dynamics. Curr Biol. 2015;25:R677–R691. - PubMed
-
- Chesarone Ma, DuPage AG, Goode BL. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat Rev Mol Cell Biol. 2010;11:62–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
