

Structural Analysis of Multi-Helical RNAs by NMR-SAXS/WAXS: Application to the U4/U6 di-snRNA
- PMID: 26655855
- PMCID: PMC4790120
- DOI: 10.1016/j.jmb.2015.11.026
Structural Analysis of Multi-Helical RNAs by NMR-SAXS/WAXS: Application to the U4/U6 di-snRNA
Abstract
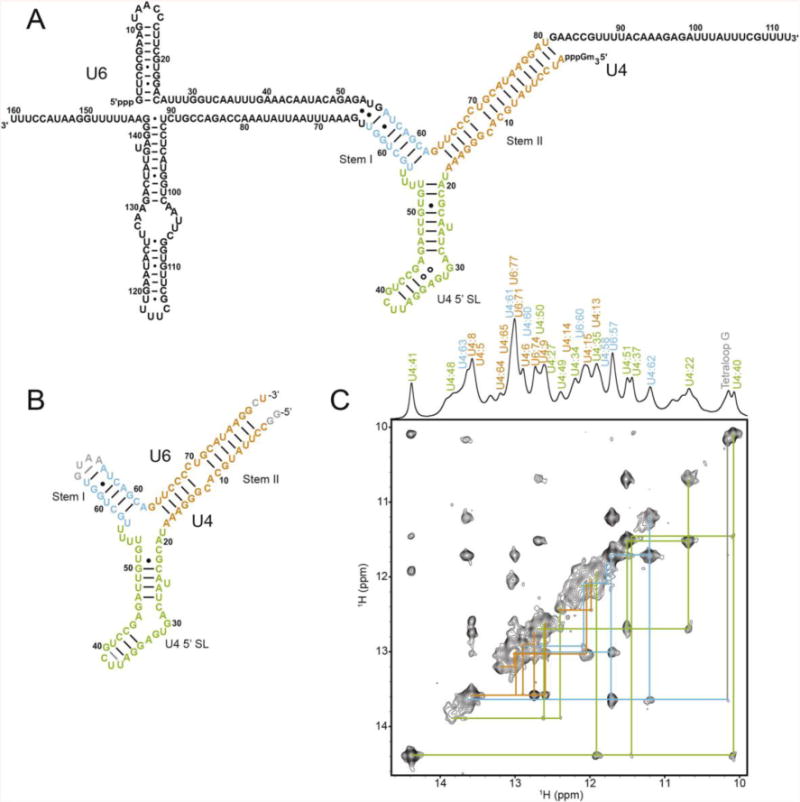
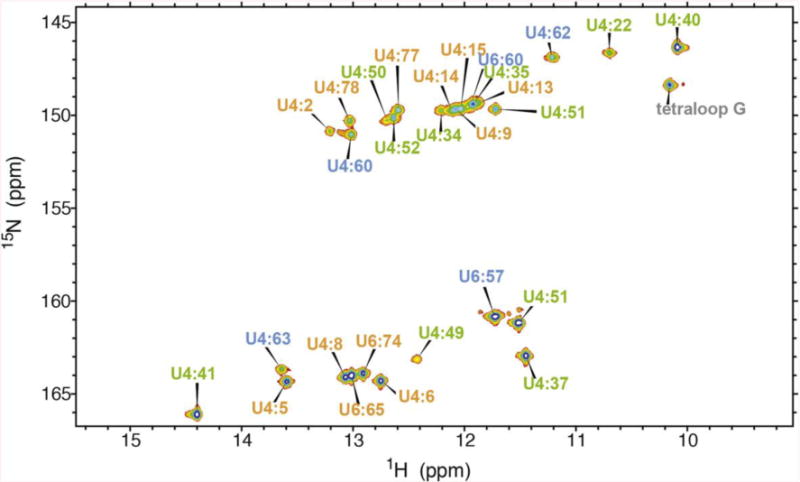
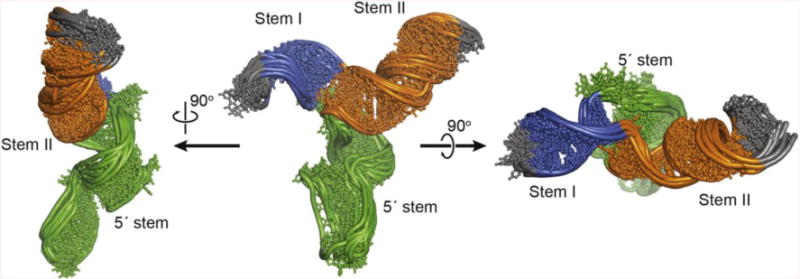
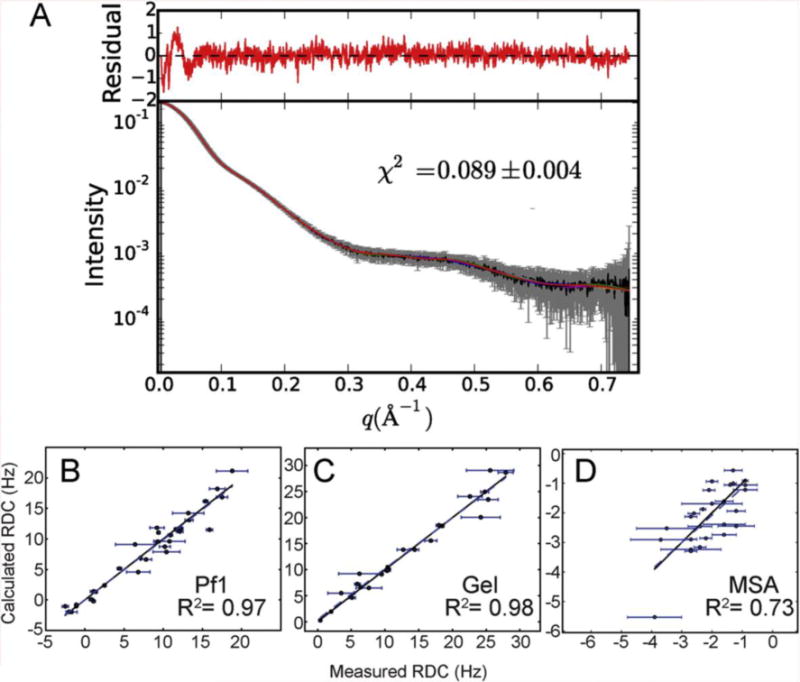
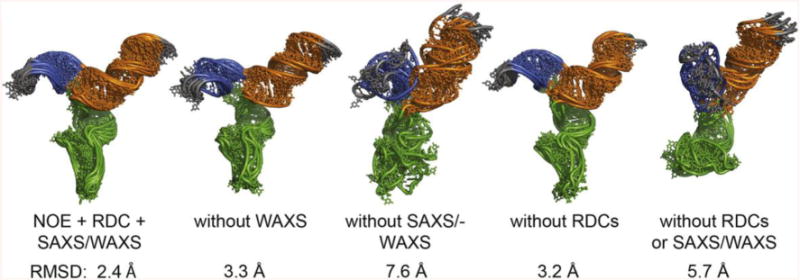
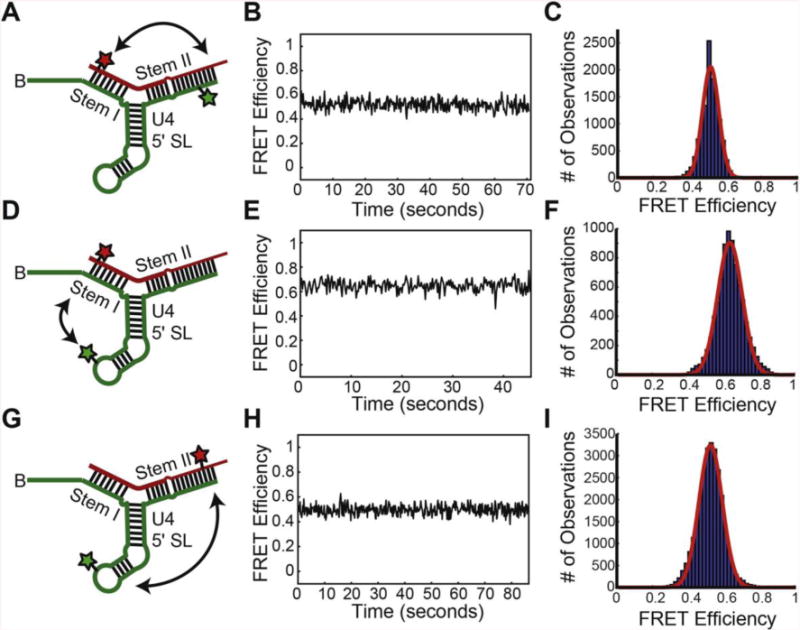
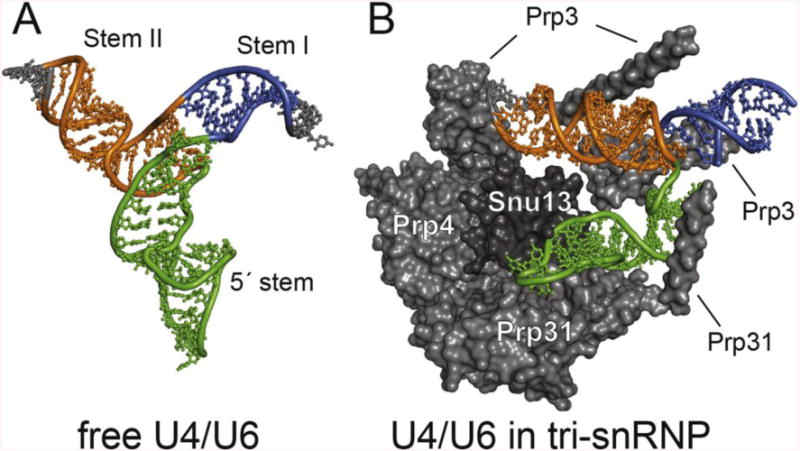
NMR and SAXS (small-angle X-ray scattering)/WAXS (wide-angle X-ray scattering) are highly complementary approaches for the analysis of RNA structure in solution. Here we describe an efficient NMR-SAXS/WAXS approach for structural investigation of multi-helical RNAs. We illustrate this approach by determining the overall fold of a 92-nt 3-helix junction from the U4/U6 di-snRNA. The U4/U6 di-snRNA is conserved in eukaryotes and is part of the U4/U6.U5 tri-snRNP, a large ribonucleoprotein complex that comprises a major subunit of the assembled spliceosome. Helical orientations can be determined by X-ray scattering data alone, but the addition of NMR RDC (residual dipolar coupling) restraints improves the structure models. RDCs were measured in two different external alignment media and also by magnetic susceptibility anisotropy. The resulting alignment tensors are collinear, which is a previously noted problem for nucleic acids. Including WAXS data in the calculations produces models with significantly better fits to the scattering data. In solution, the U4/U6 di-snRNA forms a 3-helix junction with a planar Y-shaped structure and has no detectable tertiary interactions. Single-molecule Förster resonance energy transfer data support the observed topology. A comparison with the recently determined cryo-electron microscopy structure of the U4/U6.U5 tri-snRNP illustrates how proteins scaffold the RNA and dramatically alter the geometry of the U4/U6 3-helix junction.
Keywords: NMR; RNA; SAXS; spliceosome; structure modeling.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Cate JH, Gooding AR, Podell E, Zhou K, Golden BL, Kundrot CE, et al. Crystal structure of a group I ribozyme domain: principles of RNA packing. Science. 1996;273:1678–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
