

Skp1 Independent Function of Cdc53/Cul1 in F-box Protein Homeostasis
- PMID: 26656496
- PMCID: PMC4675558
- DOI: 10.1371/journal.pgen.1005727
Skp1 Independent Function of Cdc53/Cul1 in F-box Protein Homeostasis
Abstract
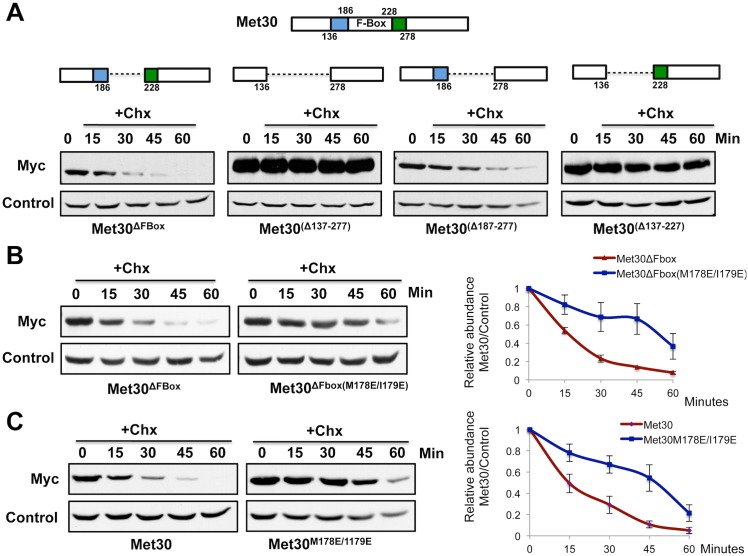
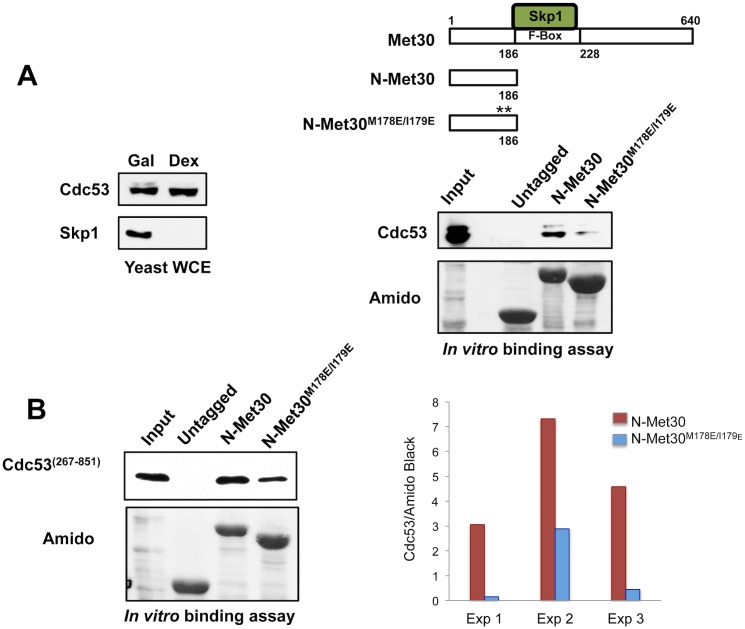
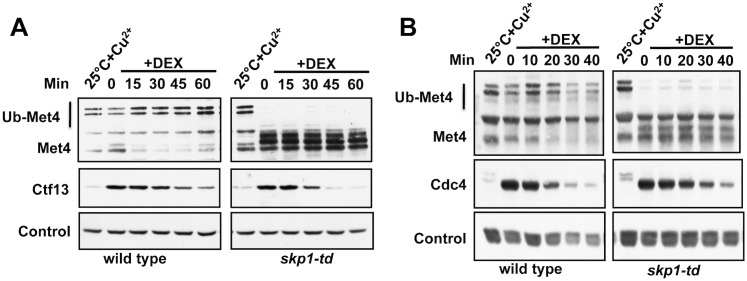
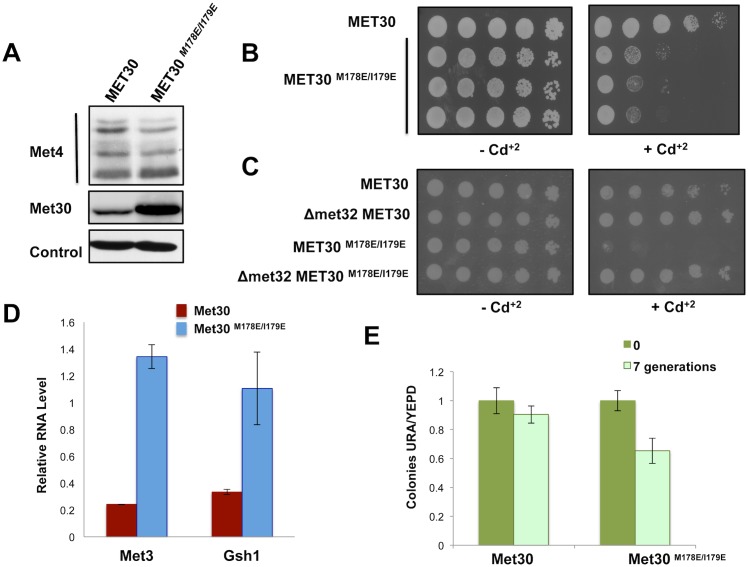
Abundance of substrate receptor subunits of Cullin-RING ubiquitin ligases (CRLs) is tightly controlled to maintain the full repertoire of CRLs. Unbalanced levels can lead to sequestration of CRL core components by a few overabundant substrate receptors. Numerous diseases, including cancer, have been associated with misregulation of substrate receptor components, particularly for the largest class of CRLs, the SCF ligases. One relevant mechanism that controls abundance of their substrate receptors, the F-box proteins, is autocatalytic ubiquitylation by intact SCF complex followed by proteasome-mediated degradation. Here we describe an additional pathway for regulation of F-box proteins on the example of yeast Met30. This ubiquitylation and degradation pathway acts on Met30 that is dissociated from Skp1. Unexpectedly, this pathway required the cullin component Cdc53/Cul1 but was independent of the other central SCF component Skp1. We demonstrated that this non-canonical degradation pathway is critical for chromosome stability and effective defense against heavy metal stress. More importantly, our results assign important biological functions to a sub-complex of cullin-RING ligases that comprises Cdc53/Rbx1/Cdc34, but is independent of Skp1.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Cdc53 is a scaffold protein for multiple Cdc34/Skp1/F-box proteincomplexes that regulate cell division and methionine biosynthesis in yeast.Genes Dev. 1998 Mar 1;12(5):692-705. doi: 10.1101/gad.12.5.692. Genes Dev. 1998. PMID: 9499404 Free PMC article.
-
SCF ubiquitin protein ligases and phosphorylation-dependent proteolysis.Philos Trans R Soc Lond B Biol Sci. 1999 Sep 29;354(1389):1533-50. doi: 10.1098/rstb.1999.0497. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10582239 Free PMC article.
-
Cdc34 and the F-box protein Met30 are required for degradation of the Cdk-inhibitory kinase Swe1.Genes Dev. 1998 Aug 15;12(16):2587-97. doi: 10.1101/gad.12.16.2587. Genes Dev. 1998. PMID: 9716410 Free PMC article.
-
Genetically engineered mouse models for functional studies of SKP1-CUL1-F-box-protein (SCF) E3 ubiquitin ligases.Cell Res. 2013 May;23(5):599-619. doi: 10.1038/cr.2013.44. Epub 2013 Mar 26. Cell Res. 2013. PMID: 23528706 Free PMC article. Review.
-
The F-box: a new motif for ubiquitin dependent proteolysis in cell cycle regulation and signal transduction.Prog Biophys Mol Biol. 1999;72(3):299-328. doi: 10.1016/s0079-6107(99)00010-3. Prog Biophys Mol Biol. 1999. PMID: 10581972 Review.
Cited by
-
F-box receptor mediated control of substrate stability and subcellular location organizes cellular development of Aspergillus nidulans.PLoS Genet. 2022 Dec 12;18(12):e1010502. doi: 10.1371/journal.pgen.1010502. eCollection 2022 Dec. PLoS Genet. 2022. PMID: 36508464 Free PMC article.
-
Cadmium binding by the F-box domain induces p97-mediated SCF complex disassembly to activate stress response programs.Nat Commun. 2024 May 8;15(1):3894. doi: 10.1038/s41467-024-48184-6. Nat Commun. 2024. PMID: 38719837 Free PMC article.
-
Cdc48 cofactor Shp1 regulates signal-induced SCFMet30 disassembly.Proc Natl Acad Sci U S A. 2020 Sep 1;117(35):21319-21327. doi: 10.1073/pnas.1922891117. Epub 2020 Aug 18. Proc Natl Acad Sci U S A. 2020. PMID: 32817489 Free PMC article.
References
-
- Hershko A, Ciechanover A (1998) The ubiquitin system. Annu Rev Biochem 67: 425–479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
