

Mutations of Conserved Residues in the Major Homology Region Arrest Assembling HIV-1 Gag as a Membrane-Targeted Intermediate Containing Genomic RNA and Cellular Proteins
- PMID: 26656702
- PMCID: PMC4734008
- DOI: 10.1128/JVI.02698-15
Mutations of Conserved Residues in the Major Homology Region Arrest Assembling HIV-1 Gag as a Membrane-Targeted Intermediate Containing Genomic RNA and Cellular Proteins
Abstract
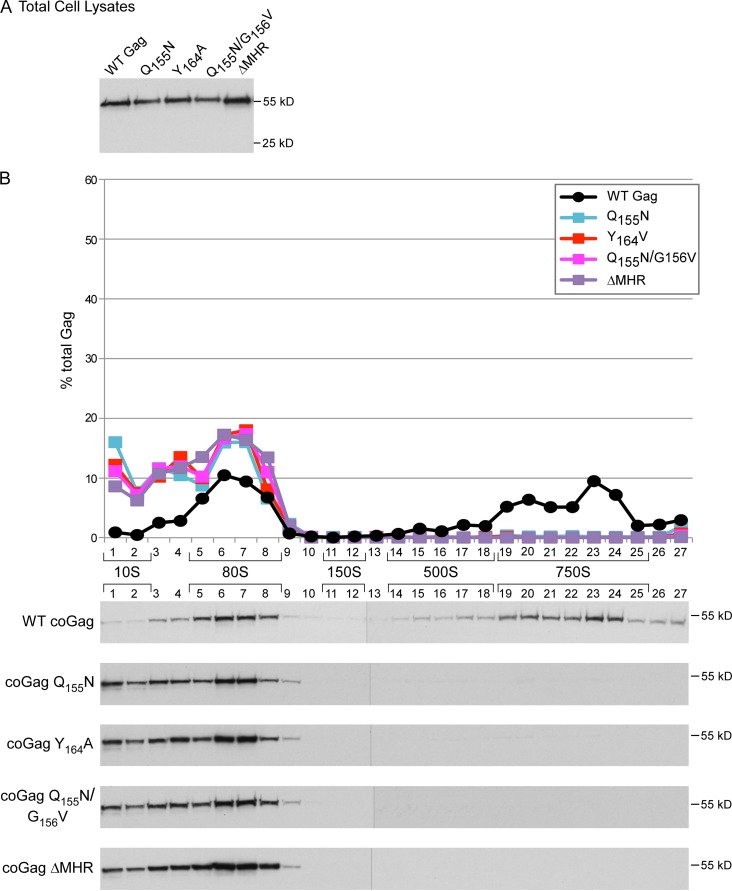
The major homology region (MHR) is a highly conserved motif that is found within the Gag protein of all orthoretroviruses and some retrotransposons. While it is widely accepted that the MHR is critical for assembly of HIV-1 and other retroviruses, how the MHR functions and why it is so highly conserved are not understood. Moreover, consensus is lacking on when HIV-1 MHR residues function during assembly. Here, we first addressed previous conflicting reports by confirming that MHR deletion, like conserved MHR residue substitution, leads to a dramatic reduction in particle production in human and nonhuman primate cells expressing HIV-1 proviruses. Next, we used biochemical analyses and immunoelectron microscopy to demonstrate that conserved residues in the MHR are required after assembling Gag has associated with genomic RNA, recruited critical host factors involved in assembly, and targeted to the plasma membrane. The exact point of inhibition at the plasma membrane differed depending on the specific mutation, with one MHR mutant arrested as a membrane-associated intermediate that is stable upon high-salt treatment and other MHR mutants arrested as labile, membrane-associated intermediates. Finally, we observed the same assembly-defective phenotypes when the MHR deletion or conserved MHR residue substitutions were engineered into Gag from a subtype B, lab-adapted provirus or Gag from a subtype C primary isolate that was codon optimized. Together, our data support a model in which MHR residues act just after membrane targeting, with some MHR residues promoting stability and another promoting multimerization of the membrane-targeted assembling Gag oligomer.
Importance: The retroviral Gag protein exhibits extensive amino acid sequence variation overall; however, one region of Gag, termed the major homology region, is conserved among all retroviruses and even some yeast retrotransposons, although the reason for this conservation remains poorly understood. Highly conserved residues in the major homology region are required for assembly of retroviruses; however, when these residues are required during assembly is not clear. Here, we used biochemical and electron microscopic analyses to demonstrate that these conserved residues function after assembling HIV-1 Gag has associated with genomic RNA, recruited critical host factors involved in assembly, and targeted to the plasma membrane but before Gag has completed the assembly process. By revealing precisely when conserved residues in the major homology region are required during assembly, these studies resolve existing controversies and set the stage for future experiments aimed at a more complete understanding of how the major homology region functions.
Copyright © 2016 Tanaka et al.
Figures











References
-
- Benit L, De Parseval N, Casella JF, Callebaut I, Cordonnier A, Heidmann T. 1997. Cloning of a new murine endogenous retrovirus, MuERV-L, with strong similarity to the human HERV-L element and with a gag coding sequence closely related to the Fv1 restriction gene. J Virol 71:5652–5657. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
