

CDK4 is an essential insulin effector in adipocytes
- PMID: 26657864
- PMCID: PMC4701556
- DOI: 10.1172/JCI81480
CDK4 is an essential insulin effector in adipocytes
Erratum in
-
CDK4 is an essential insulin effector in adipocytes.J Clin Invest. 2022 Jul 1;132(13):e162359. doi: 10.1172/JCI162359. J Clin Invest. 2022. PMID: 35775487 Free PMC article. No abstract available.
Retraction in
-
CDK4 is an essential insulin effector in adipocytes.J Clin Invest. 2023 Mar 15;133(6):e170315. doi: 10.1172/JCI170315. J Clin Invest. 2023. PMID: 36919703 Free PMC article. No abstract available.
Abstract
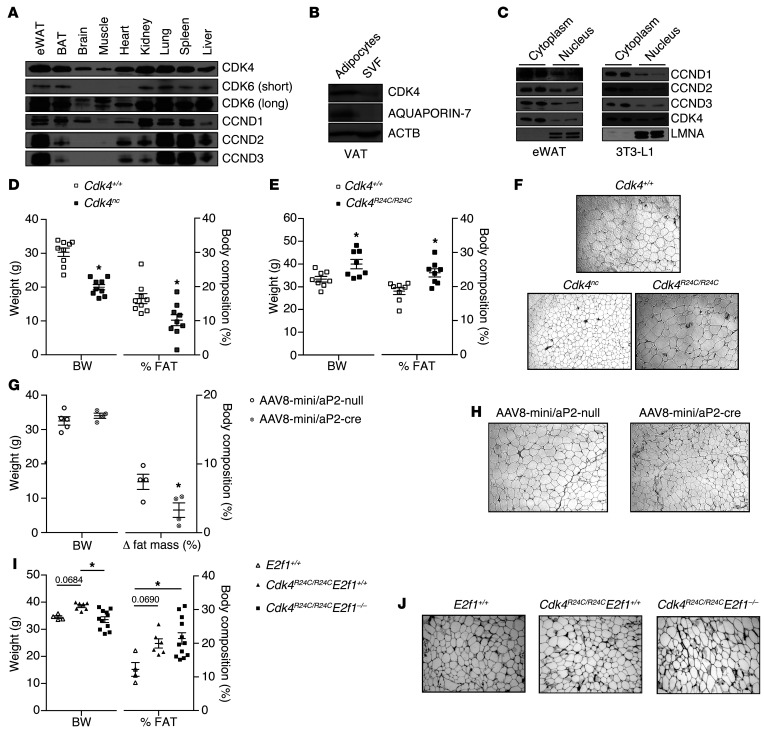
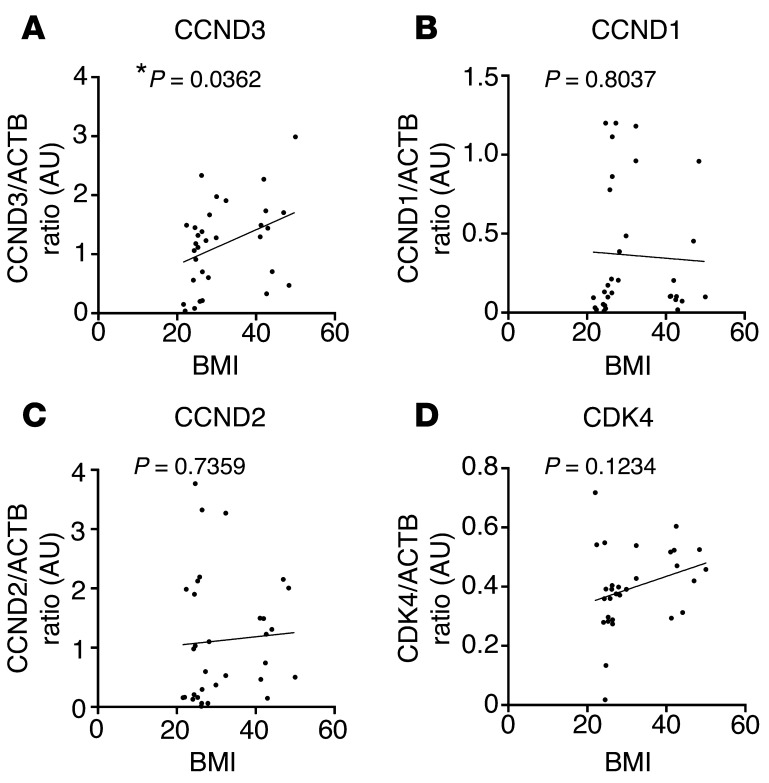
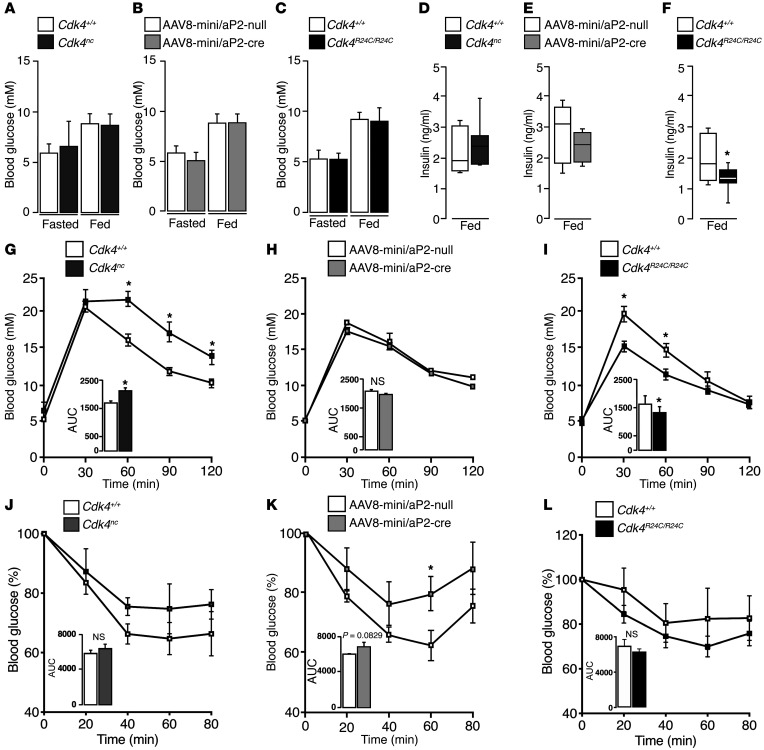
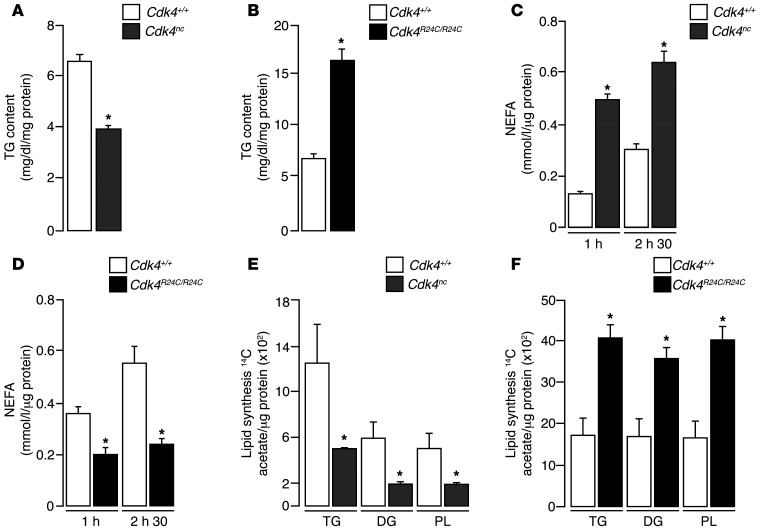
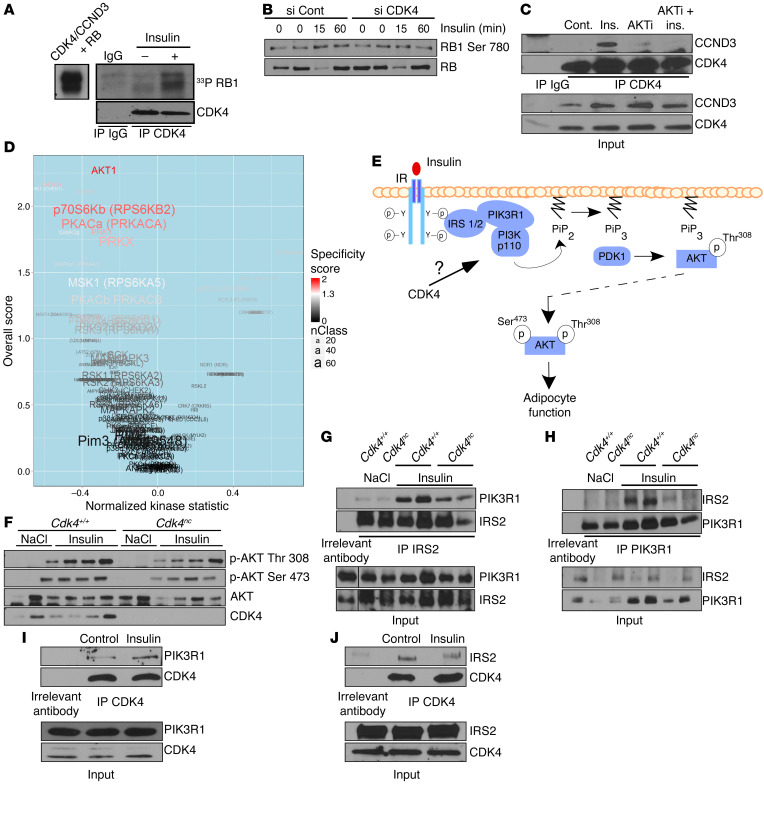
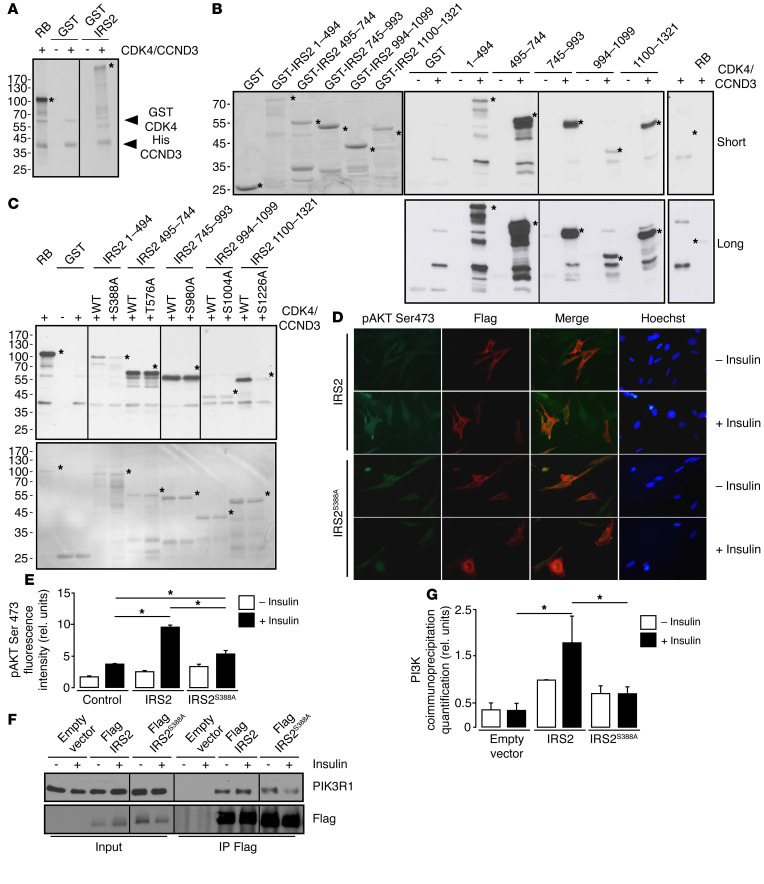
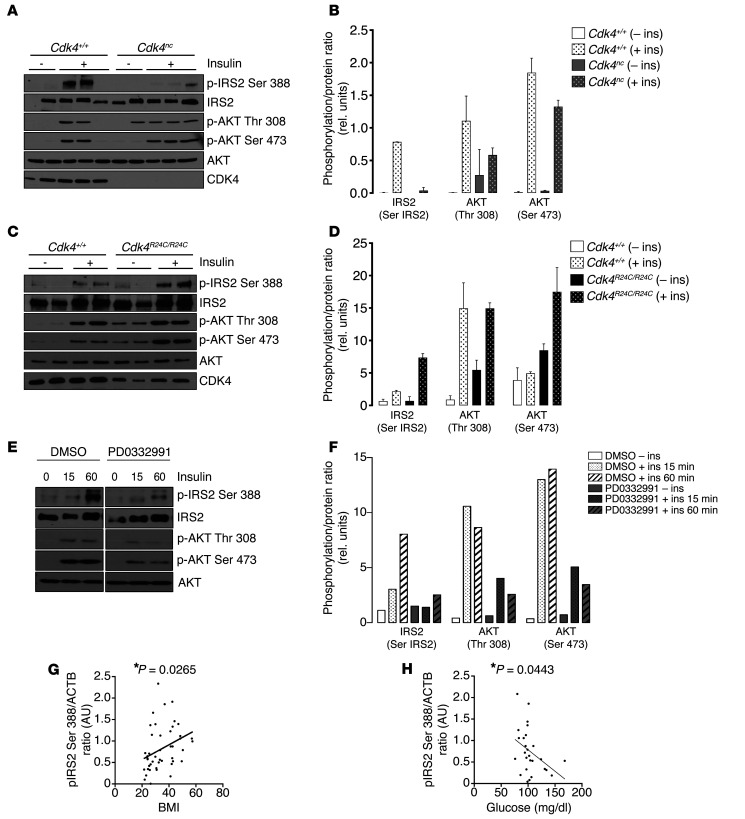
Insulin resistance is a fundamental pathogenic factor that characterizes various metabolic disorders, including obesity and type 2 diabetes. Adipose tissue contributes to the development of obesity-related insulin resistance through increased release of fatty acids, altered adipokine secretion, and/or macrophage infiltration and cytokine release. Here, we aimed to analyze the participation of the cyclin-dependent kinase 4 (CDK4) in adipose tissue biology. We determined that white adipose tissue (WAT) from CDK4-deficient mice exhibits impaired lipogenesis and increased lipolysis. Conversely, lipolysis was decreased and lipogenesis was increased in mice expressing a mutant hyperactive form of CDK4 (CDK4(R24C)). A global kinome analysis of CDK4-deficient mice following insulin stimulation revealed that insulin signaling is impaired in these animals. We determined that insulin activates the CCND3-CDK4 complex, which in turn phosphorylates insulin receptor substrate 2 (IRS2) at serine 388, thereby creating a positive feedback loop that maintains adipocyte insulin signaling. Furthermore, we found that CCND3 expression and IRS2 serine 388 phosphorylation are increased in human obese subjects. Together, our results demonstrate that CDK4 is a major regulator of insulin signaling in WAT.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
