

Respiratory Syncytial Virus Uses CX3CR1 as a Receptor on Primary Human Airway Epithelial Cultures
- PMID: 26658574
- PMCID: PMC4676609
- DOI: 10.1371/journal.ppat.1005318
Respiratory Syncytial Virus Uses CX3CR1 as a Receptor on Primary Human Airway Epithelial Cultures
Abstract
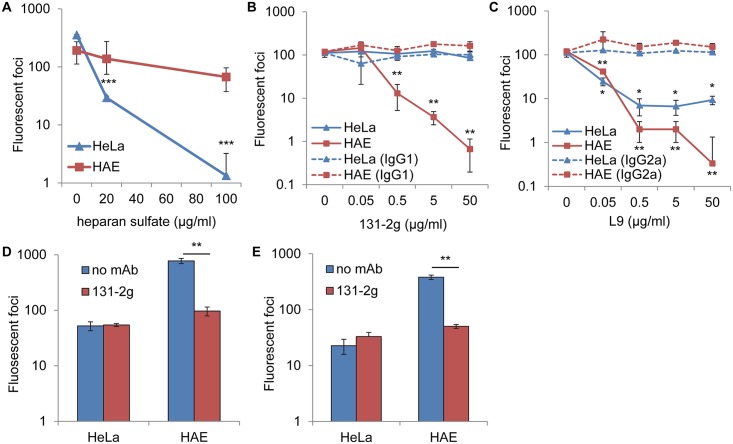
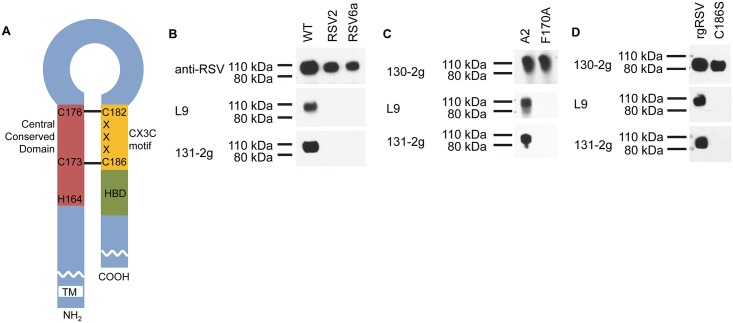
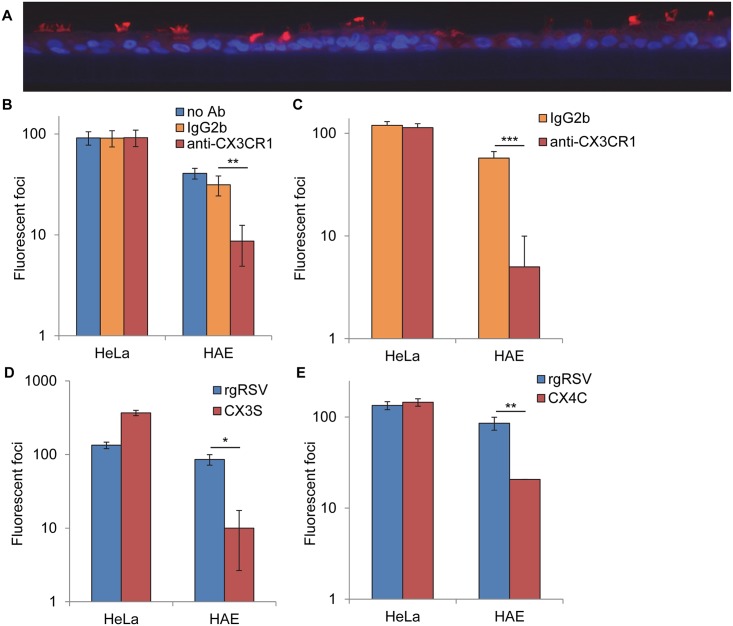
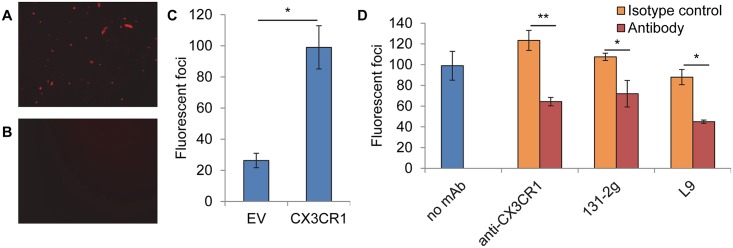
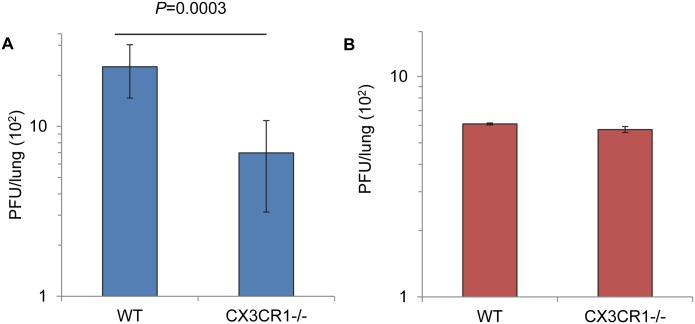
Respiratory syncytial virus (RSV) is the most frequent cause of lower respiratory disease in infants, but no vaccine or effective therapy is available. The initiation of RSV infection of immortalized cells is largely dependent on cell surface heparan sulfate (HS), a receptor for the RSV attachment (G) glycoprotein in immortalized cells. However, RSV infects the ciliated cells in primary well differentiated human airway epithelial (HAE) cultures via the apical surface, but HS is not detectable on this surface. Here we show that soluble HS inhibits infection of immortalized cells, but not HAE cultures, confirming that HS is not the receptor on HAE cultures. Conversely, a "non-neutralizing" monoclonal antibody against the G protein that does not block RSV infection of immortalized cells, does inhibit infection of HAE cultures. This antibody was previously shown to block the interaction between the G protein and the chemokine receptor CX3CR1 and we have mapped the binding site for this antibody to the CX3C motif and its surrounding region in the G protein. We show that CX3CR1 is present on the apical surface of ciliated cells in HAE cultures and especially on the cilia. RSV infection of HAE cultures is reduced by an antibody against CX3CR1 and by mutations in the G protein CX3C motif. Additionally, mice lacking CX3CR1 are less susceptible to RSV infection. These findings demonstrate that RSV uses CX3CR1 as a cellular receptor on HAE cultures and highlight the importance of using a physiologically relevant model to study virus entry and antibody neutralization.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Glezen WP, Taber LH, Frank AL, Kasel JA (1986) Risk of primary infection and reinfection with respiratory syncytial virus. Am J Dis Child 140: 543–546. - PubMed
-
- Holberg CJ, Wright AL, Martinez FD, Ray CG, Taussig LM, et al. (1991) Risk factors for respiratory syncytial virus-associated lower respiratory illnesses in the first year of life. Am J Epidemiol 133: 1135–1151. - PubMed
-
- Langley JM, LeBlanc JC, Smith B, Wang EE (2003) Increasing incidence of hospitalization for bronchiolitis among Canadian children, 1980–2000. J Infect Dis 188: 1764–1767. - PubMed
-
- Shay DK, Holman RC, Newman RD, Liu LL, Stout JW, et al. (1999) Bronchiolitis-associated hospitalizations among US children, 1980–1996. JAMA 282: 1440–1446. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
