

A Unified Dynamic Model for Learning, Replay, and Sharp-Wave/Ripples
- PMID: 26658873
- PMCID: PMC6605500
- DOI: 10.1523/JNEUROSCI.3977-14.2015
A Unified Dynamic Model for Learning, Replay, and Sharp-Wave/Ripples
Abstract
Hippocampal activity is fundamental for episodic memory formation and consolidation. During phases of rest and sleep, it exhibits sharp-wave/ripple (SPW/R) complexes, which are short episodes of increased activity with superimposed high-frequency oscillations. Simultaneously, spike sequences reflecting previous behavior, such as traversed trajectories in space, are replayed. Whereas these phenomena are thought to be crucial for the formation and consolidation of episodic memory, their neurophysiological mechanisms are not well understood. Here we present a unified model showing how experience may be stored and thereafter replayed in association with SPW/Rs. We propose that replay and SPW/Rs are tightly interconnected as they mutually generate and support each other. The underlying mechanism is based on the nonlinear dendritic computation attributable to dendritic sodium spikes that have been prominently found in the hippocampal regions CA1 and CA3, where SPW/Rs and replay are also generated. Besides assigning SPW/Rs a crucial role for replay and thus memory processing, the proposed mechanism also explains their characteristic features, such as the oscillation frequency and the overall wave form. The results shed a new light on the dynamical aspects of hippocampal circuit learning.
Significance statement: During phases of rest and sleep, the hippocampus, the "memory center" of the brain, generates intermittent patterns of strongly increased overall activity with high-frequency oscillations, the so-called sharp-wave/ripples. We investigate their role in learning and memory processing. They occur together with replay of activity sequences reflecting previous behavior. Developing a unifying computational model, we propose that both phenomena are tightly linked, by mutually generating and supporting each other. The underlying mechanism depends on nonlinear amplification of synchronous inputs that has been prominently found in the hippocampus. Besides assigning sharp-wave/ripples a crucial role for replay generation and thus memory processing, the proposed mechanism also explains their characteristic features, such as the oscillation frequency and the overall wave form.
Keywords: dendritic spikes; learning; memory; network; replay; sharp-wave/ripples.
Copyright © 2015 the authors 0270-6474/15/3516236-23$15.00/0.
Figures
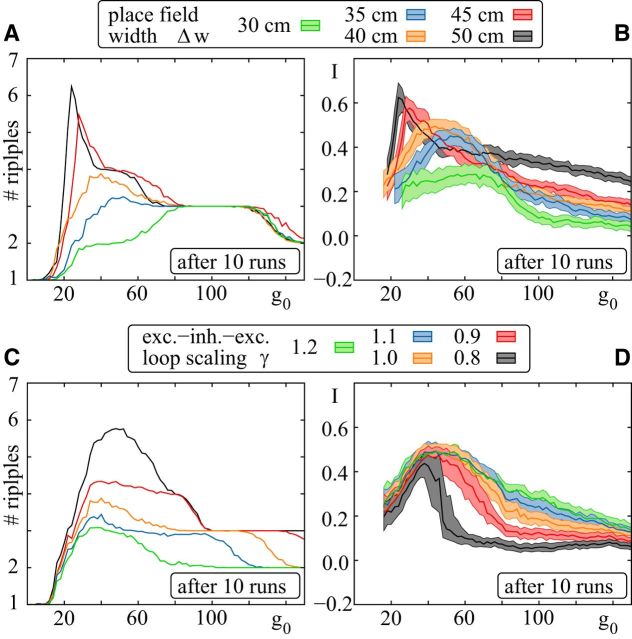
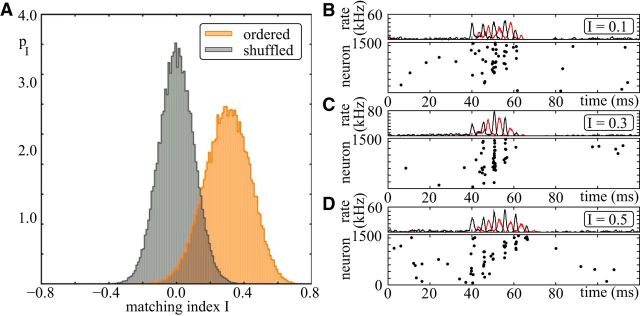
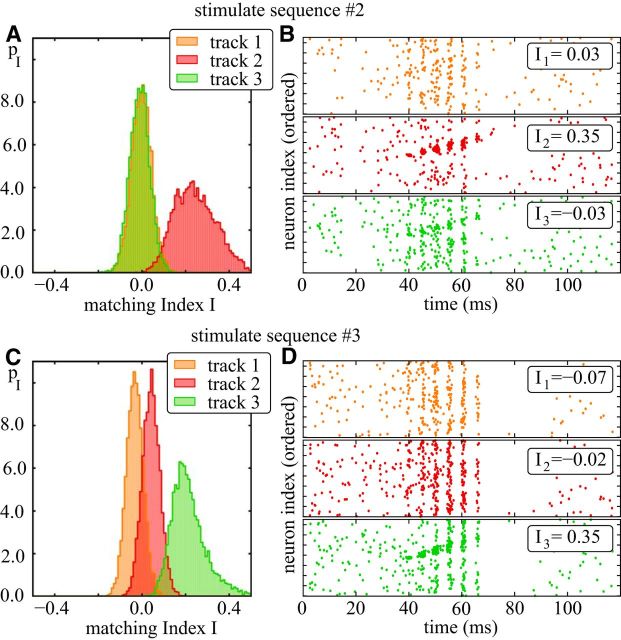
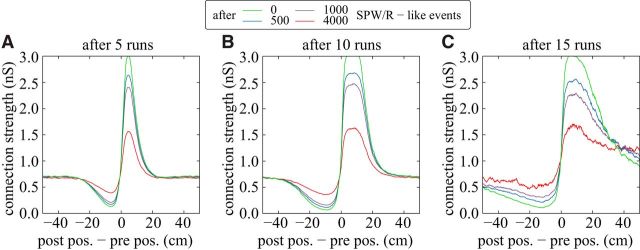
Comment in
-
Dendritic Spikes Provide a Mechanism for Hippocampal Replay and Sharp-Wave/Ripple Generation.J Neurosci. 2016 Apr 13;36(15):4152-4. doi: 10.1523/JNEUROSCI.0442-16.2016. J Neurosci. 2016. PMID: 27076415 Free PMC article. No abstract available.
Similar articles
-
Impairment of Sharp-Wave Ripples in a Murine Model of Dravet Syndrome.J Neurosci. 2019 Nov 13;39(46):9251-9260. doi: 10.1523/JNEUROSCI.0890-19.2019. Epub 2019 Sep 19. J Neurosci. 2019. PMID: 31537705 Free PMC article.
-
Origin of Gamma Frequency Power during Hippocampal Sharp-Wave Ripples.Cell Rep. 2018 Nov 13;25(7):1693-1700.e4. doi: 10.1016/j.celrep.2018.10.066. Cell Rep. 2018. PMID: 30428340 Free PMC article.
-
Most hippocampal CA1 pyramidal cells in rabbits increase firing during awake sharp-wave ripples and some do so in response to external stimulation and theta.J Neurophysiol. 2020 May 1;123(5):1671-1681. doi: 10.1152/jn.00056.2020. Epub 2020 Mar 25. J Neurophysiol. 2020. PMID: 32208887
-
Hippocampal sharp wave-ripple: A cognitive biomarker for episodic memory and planning.Hippocampus. 2015 Oct;25(10):1073-188. doi: 10.1002/hipo.22488. Hippocampus. 2015. PMID: 26135716 Free PMC article. Review.
-
Hippocampal ripples as a mode of communication with cortical and subcortical areas.Hippocampus. 2020 Jan;30(1):39-49. doi: 10.1002/hipo.22997. Epub 2018 Nov 13. Hippocampus. 2020. PMID: 30069976 Review.
Cited by
-
When less is more: non-monotonic spike sequence processing in neurons.PLoS Comput Biol. 2015 Feb 3;11(2):e1004002. doi: 10.1371/journal.pcbi.1004002. eCollection 2015 Feb. PLoS Comput Biol. 2015. PMID: 25646860 Free PMC article.
-
Differences in the consolidation by spontaneous and evoked ripples in the presence of active dendrites.PLoS Comput Biol. 2024 Jun 25;20(6):e1012218. doi: 10.1371/journal.pcbi.1012218. eCollection 2024 Jun. PLoS Comput Biol. 2024. PMID: 38917228 Free PMC article.
-
Emergent structural and functional properties of hippocampal multi-cellular aggregates.Front Neurosci. 2023 Jun 15;17:1171115. doi: 10.3389/fnins.2023.1171115. eCollection 2023. Front Neurosci. 2023. PMID: 37397454 Free PMC article.
-
Storage fidelity for sequence memory in the hippocampal circuit.PLoS One. 2018 Oct 4;13(10):e0204685. doi: 10.1371/journal.pone.0204685. eCollection 2018. PLoS One. 2018. PMID: 30286147 Free PMC article.
-
A neural network model for online one-shot storage of pattern sequences.PLoS One. 2024 Jun 20;19(6):e0304076. doi: 10.1371/journal.pone.0304076. eCollection 2024. PLoS One. 2024. PMID: 38900733 Free PMC article.
References
-
- Abeles M. Local cortical circuits: an electrophysiological study. Berlin: Springer; 1982.
-
- Abeles M. Corticonics: neural circuits of the cerebral cortex. Cambridge, UK: Cambridge UP; 1991.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous