

Network mechanisms of intentional learning
- PMID: 26658925
- PMCID: PMC4758826
- DOI: 10.1016/j.neuroimage.2015.11.060
Network mechanisms of intentional learning
Abstract
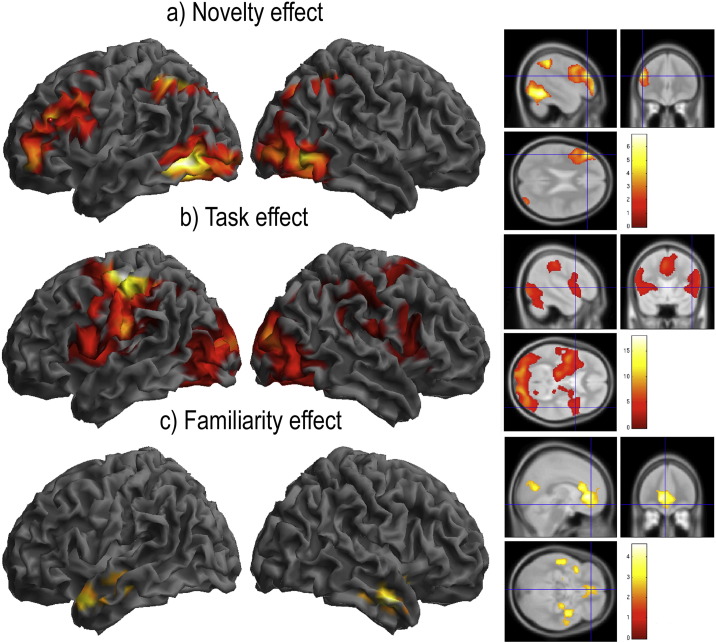
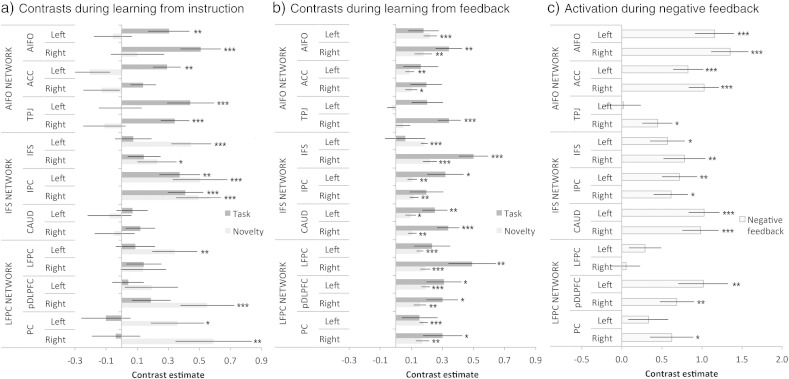
The ability to learn new tasks rapidly is a prominent characteristic of human behaviour. This ability relies on flexible cognitive systems that adapt in order to encode temporary programs for processing non-automated tasks. Previous functional imaging studies have revealed distinct roles for the lateral frontal cortices (LFCs) and the ventral striatum in intentional learning processes. However, the human LFCs are complex; they house multiple distinct sub-regions, each of which co-activates with a different functional network. It remains unclear how these LFC networks differ in their functions and how they coordinate with each other, and the ventral striatum, to support intentional learning. Here, we apply a suite of fMRI connectivity methods to determine how LFC networks activate and interact at different stages of two novel tasks, in which arbitrary stimulus-response rules are learnt either from explicit instruction or by trial-and-error. We report that the networks activate en masse and in synchrony when novel rules are being learnt from instruction. However, these networks are not homogeneous in their functions; instead, the directed connectivities between them vary asymmetrically across the learning timecourse and they disengage from the task sequentially along a rostro-caudal axis. Furthermore, when negative feedback indicates the need to switch to alternative stimulus-response rules, there is additional input to the LFC networks from the ventral striatum. These results support the hypotheses that LFC networks interact as a hierarchical system during intentional learning and that signals from the ventral striatum have a driving influence on this system when the internal program for processing the task is updated.
Keywords: Caudate; Dynamic causal modelling; Frontal cortex; Functional connectivity; Learning.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures









References
-
- Badre D. Cognitive control, hierarchy, and the rostro-caudal organization of the frontal lobes. Trends Cogn. Sci. 2008;12:193–200. - PubMed
-
- Beckmann C.F., Smith S.M. Probabilistic independent component analysis for functional magnetic resonance imaging. IEEE Trans. Med. Imaging. 2004;23:137–152. - PubMed
-
- Brett M., Anton J., Valabregue R., Poline J. 8th International Conference on Functional Mapping of the Human Brain, Sendai, Japan. 2002. Region of interest analysis using an SPM toolbox [abstract]
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
