

The dawn of active genetics
- PMID: 26660392
- PMCID: PMC5819344
- DOI: 10.1002/bies.201500102
The dawn of active genetics
Abstract
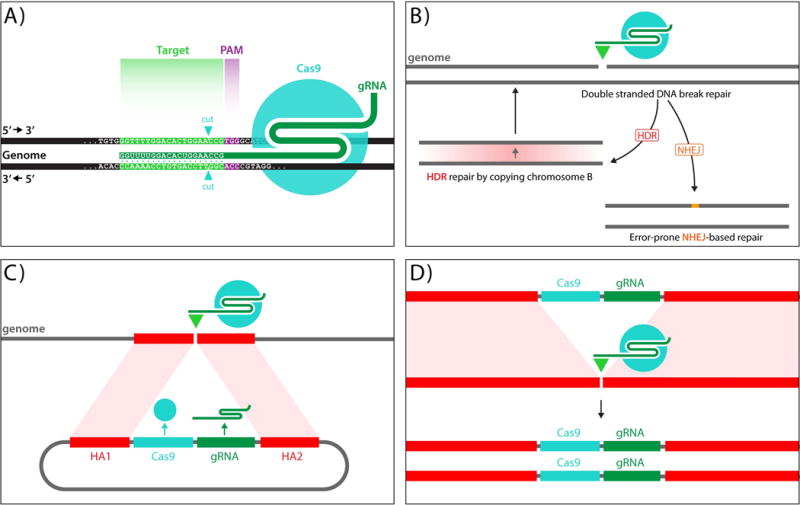
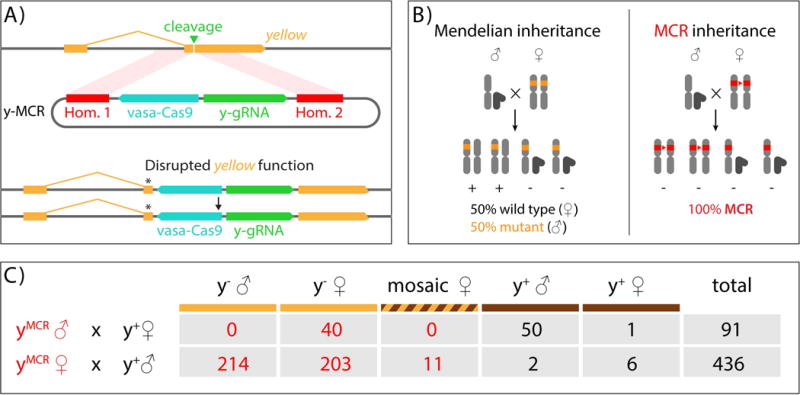
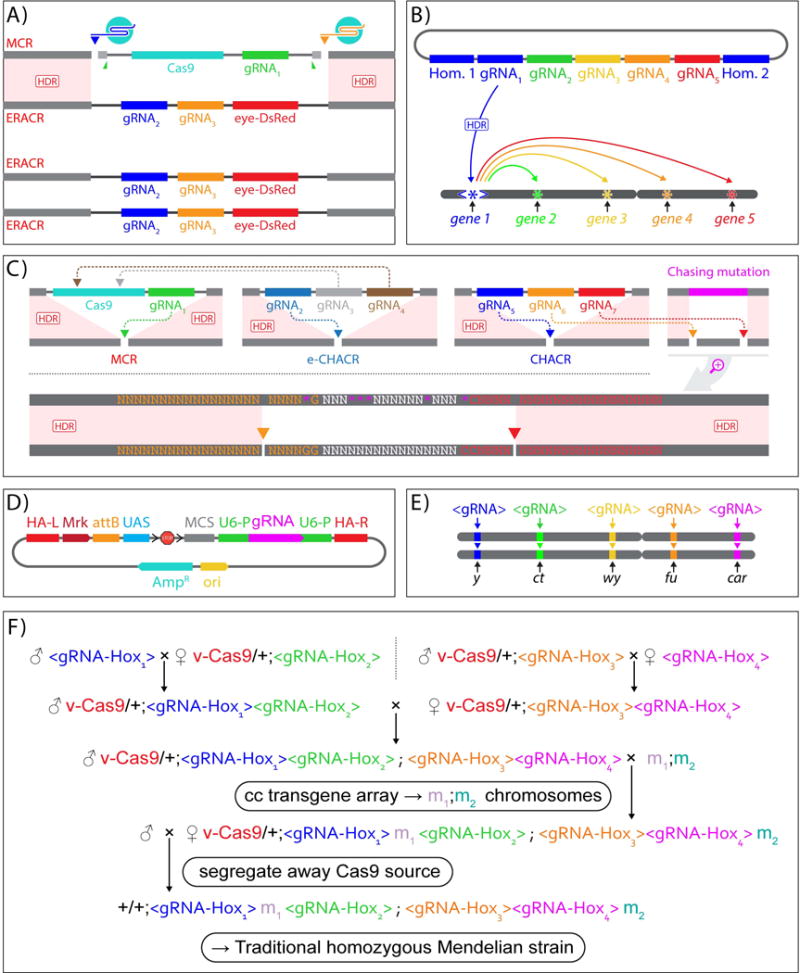
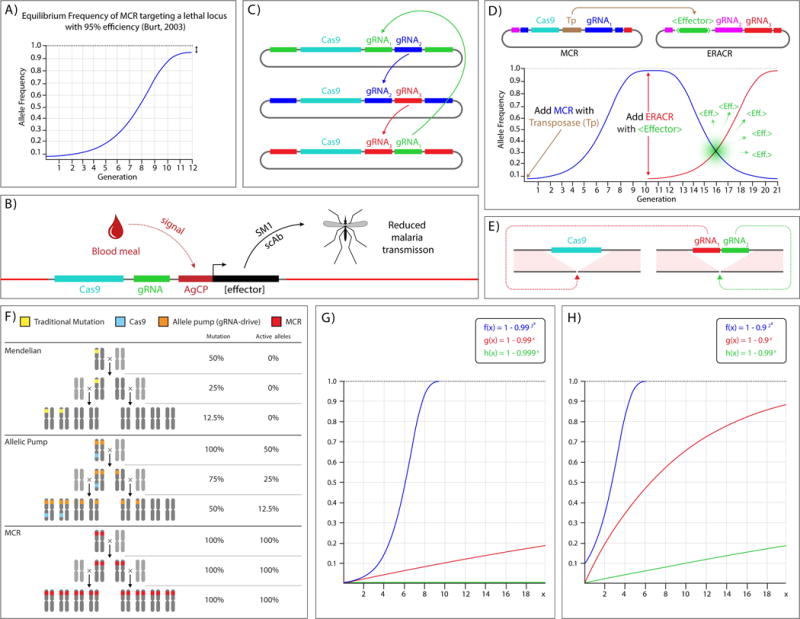
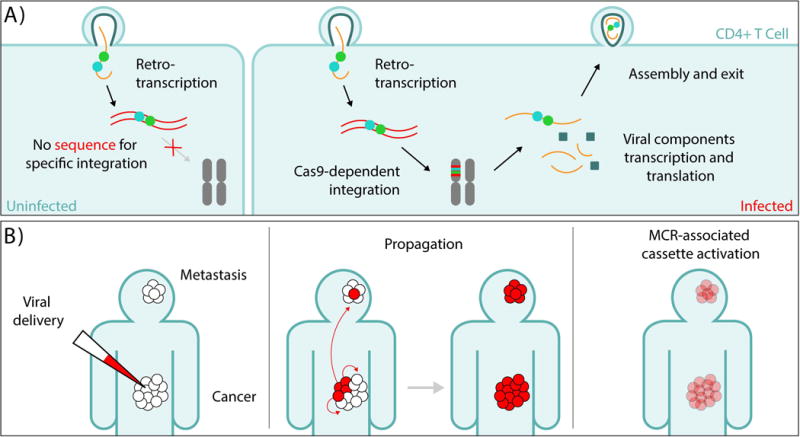
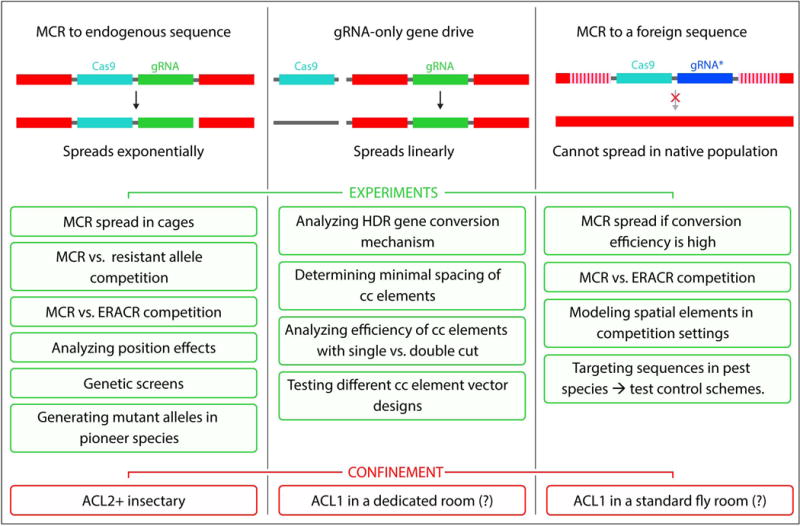
On December 18, 2014, a yellow female fly quietly emerged from her pupal case. What made her unique was that she had only one parent carrying a mutant allele of this classic recessive locus. Then, one generation later, after mating with a wild-type male, all her offspring displayed the same recessive yellow phenotype. Further analysis of other such yellow females revealed that the construct causing the mutation was converting the opposing chromosome with 95% efficiency. These simple results, seen also in mosquitoes and yeast, open the door to a new era of genetics wherein the laws of traditional Mendelian inheritance can be bypassed for a broad variety of purposes. Here, we consider the implications of this fundamentally new form of "active genetics," its applications for gene drives, reversal and amplification strategies, its potential for contributing to cell and gene therapy strategies, and ethical/biosafety considerations associated with such active genetic elements. Also watch the Video Abstract.
Keywords: Drosophila; ERACR; MCR; active genetics; copy cat element; gene drive; mutagenic chain reaction.
© 2015 WILEY Periodicals, Inc.
Figures
References
-
- Barrangou R. The roles of CRISPR-Cas systems in adaptive immunity and beyond. Curr Opin Immunol. 2015;32C:36–41. - PubMed
-
- Westra ER, Buckling A, Fineran PC. CRISPR-Cas systems: beyond adaptive immunity. Nat Rev Microbiol. 2014;12(5):317–26. - PubMed
-
- Bassett AR, Liu JL. CRISPR/Cas9 and genome editing in Drosophila. J Genet Genomics. 2014;41(1):7–19. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
