

Co-regulation of mitochondrial respiration by proline dehydrogenase/oxidase and succinate
- PMID: 26660760
- PMCID: PMC4752965
- DOI: 10.1007/s00726-015-2134-7
Co-regulation of mitochondrial respiration by proline dehydrogenase/oxidase and succinate
Erratum in
-
Erratum to: Co-regulation of mitochondrial respiration by proline dehydrogenase/oxidase and succinate.Amino Acids. 2016 May;48(5):1337-8. doi: 10.1007/s00726-016-2184-5. Amino Acids. 2016. PMID: 26874699 Free PMC article. No abstract available.
Abstract
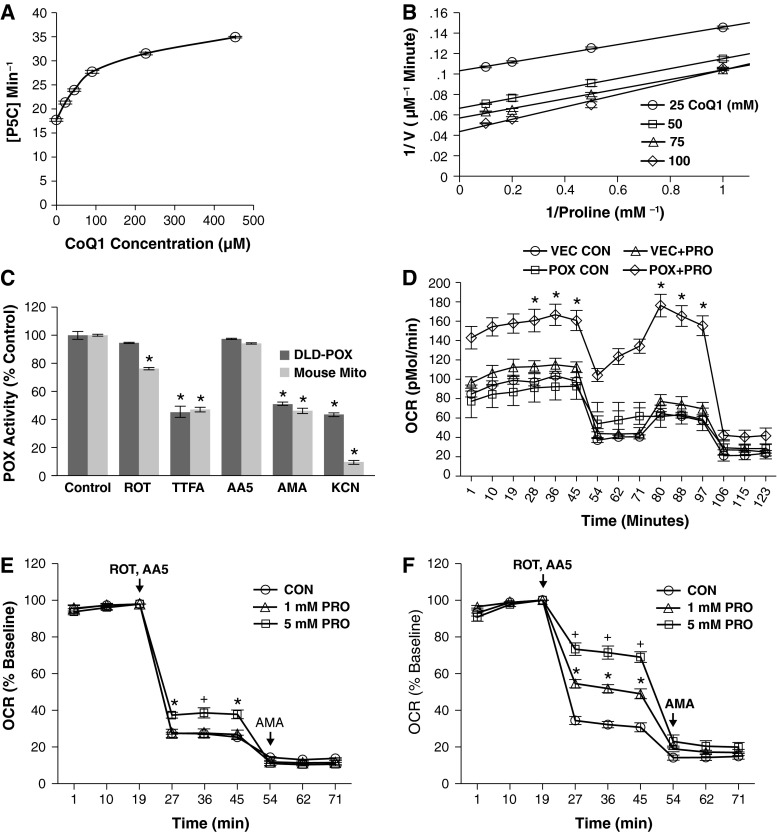
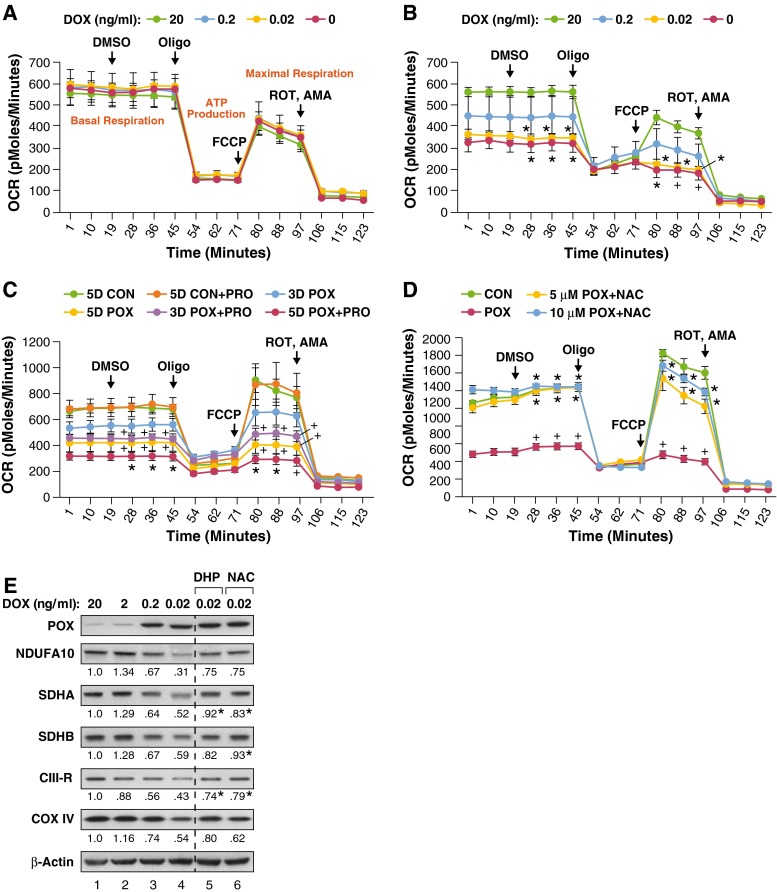
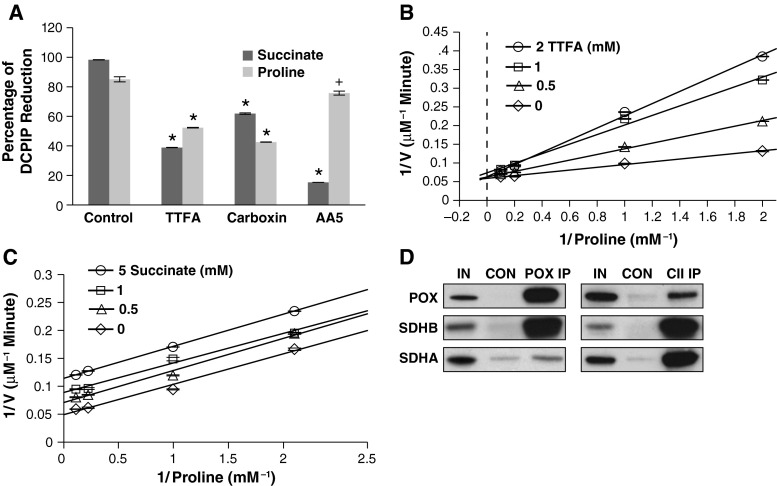
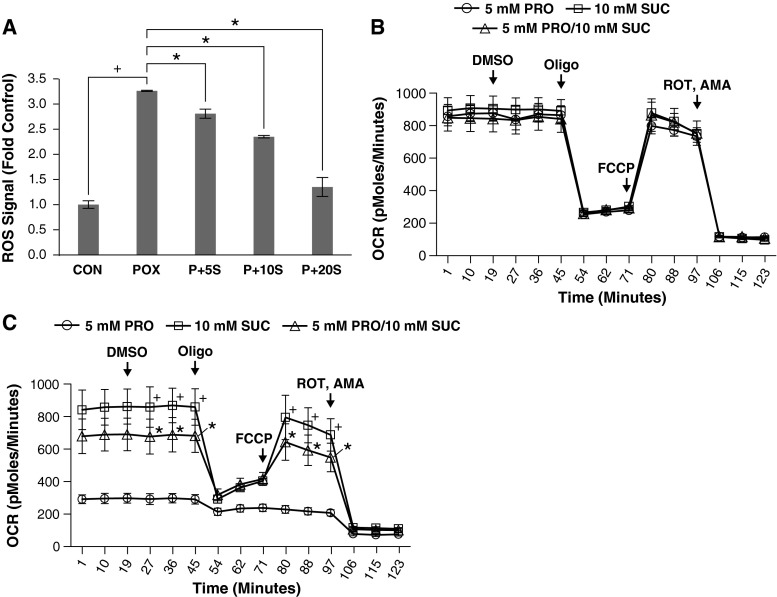
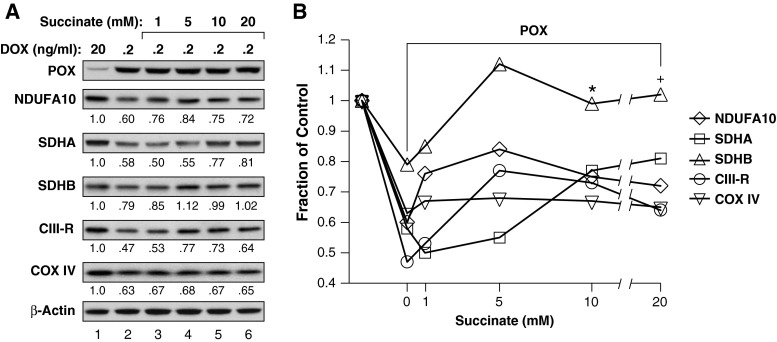
Proline dehydrogenase/oxidase (PRODH/POX) is a mitochondrial protein critical to multiple stress pathways. Because of the roles of PRODH/POX in signaling, and its shared localization to the mitochondrial inner membrane with the electron transport chain (ETC), we investigated whether there was a direct relationship between PRODH/POX and regulation of the ETC. We found that PRODH/POX binds directly to CoQ1 and that CoQ1-dependent PRODH/POX activity required functional Complex III and Complex IV. PRODH/POX supported respiration in living cells during nutrient stress; however, expression of PRODH/POX resulted in an overall decrease in respiratory fitness. Effects on respiratory fitness were inhibited by DHP and NAC, indicating that these effects were mediated by PRODH/POX-dependent reactive oxygen species (ROS) generation. PRODH/POX expression resulted in a dose-dependent down-regulation of Complexes I-IV of the ETC, and this effect was also mitigated by the addition of DHP and NAC. We found that succinate was an uncompetitive inhibitor of PRODH/POX activity, inhibited ROS generation by PRODH/POX, and alleviated PRODH/POX effects on respiratory fitness. The findings demonstrate novel cross-talk between proline and succinate respiration in vivo and provide mechanistic insights into observations from previous animal studies. Our results suggest a potential regulatory loop between PRODH/POX and succinate in regulation of mitochondrial respiration.
Keywords: Energetics; Reactive oxygen species; Redox; Respiration; Signaling.
Figures
References
-
- Alvord W (2014) Introduction to sla pdf. http://cran.r-project.org/web/packages/sla/vignettes/introduction-to-sla...
-
- Astuti D, Latif F, Dallol A, Dahia PL, Douglas F, George E, Skoldberg F, Husebye ES, Eng C, Maher ER. Gene mutations in the succinate dehydrogenase subunit SDHB cause susceptibility to familial pheochromocytoma and to familial paraganglioma. Am J Hum Genet. 2001;69(1):49–54. doi: 10.1086/321282. - DOI - PMC - PubMed
-
- N AWaC (2014) sla: two-group striaght line ANCOVA. R package version 0.1. http://CRAN.R-project.org/package=sla
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
