

Extracellular HCO3- is sensed by mouse cerebral arteries: Regulation of tone by receptor protein tyrosine phosphatase γ
- PMID: 26661205
- PMCID: PMC4853837
- DOI: 10.1177/0271678X15610787
Extracellular HCO3- is sensed by mouse cerebral arteries: Regulation of tone by receptor protein tyrosine phosphatase γ
Abstract
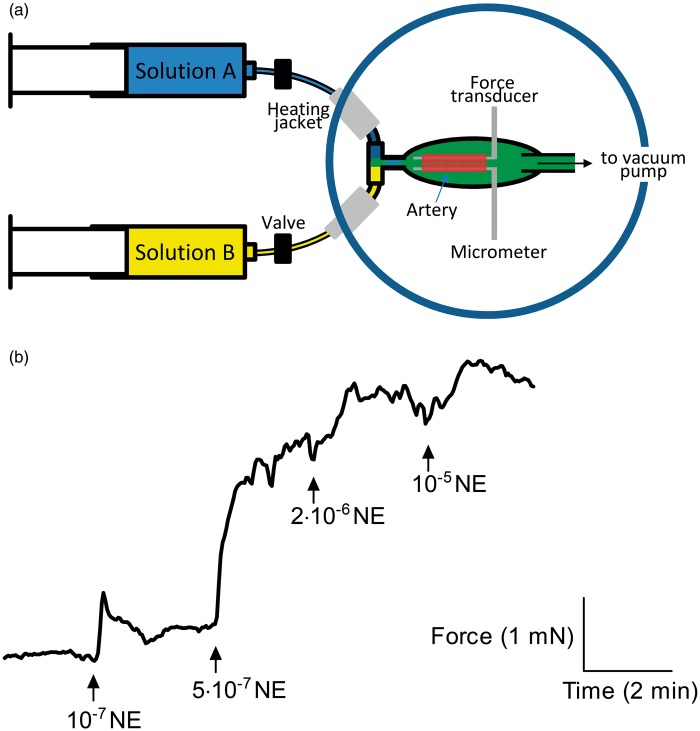
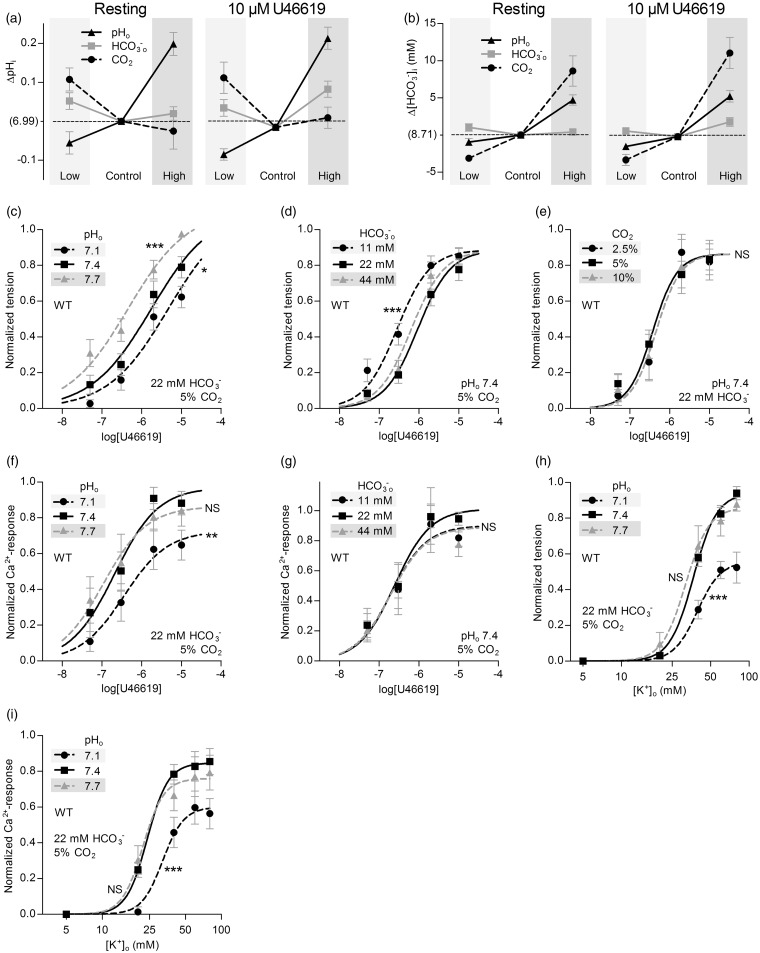
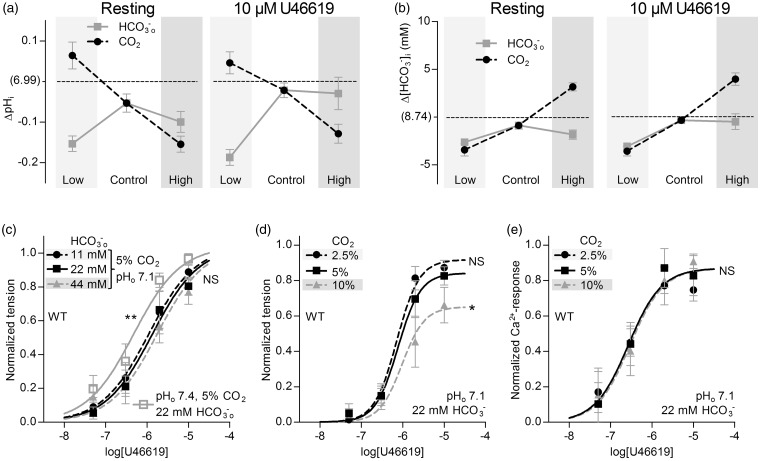
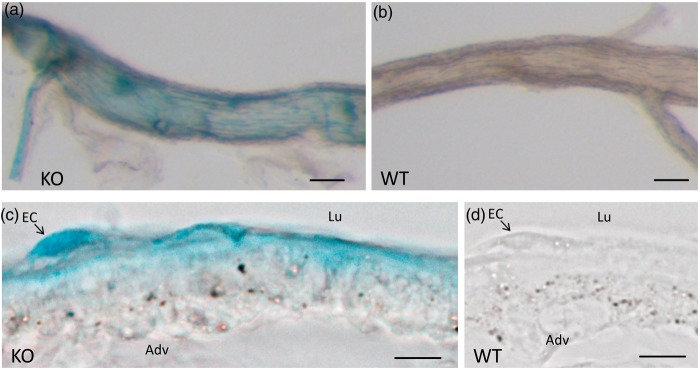
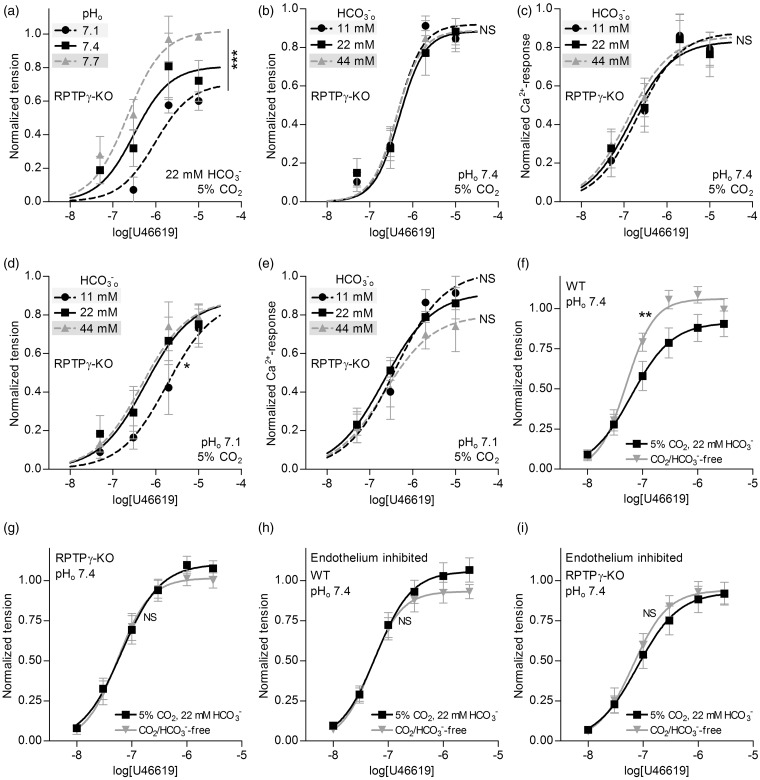
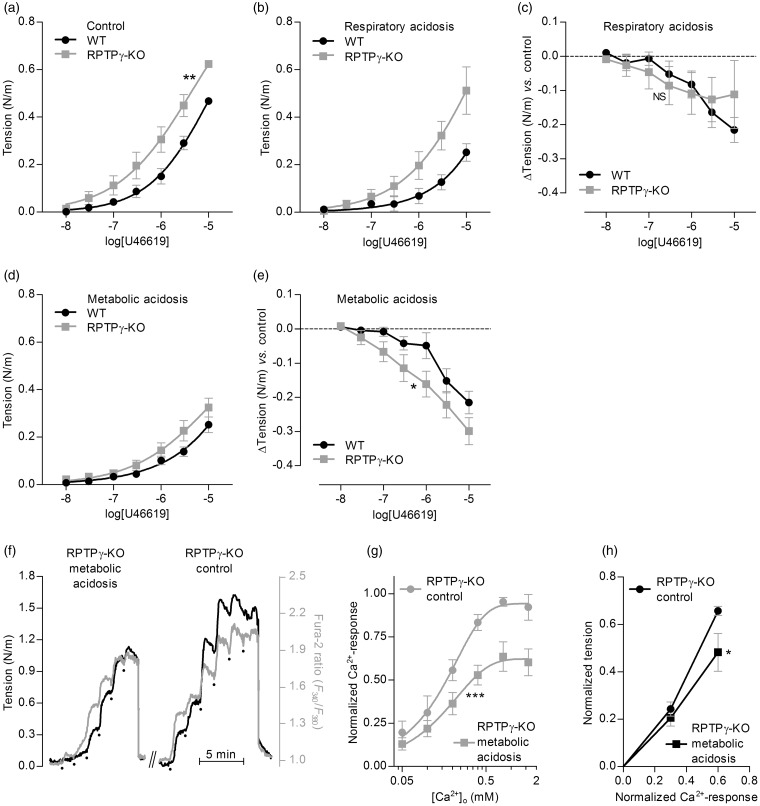
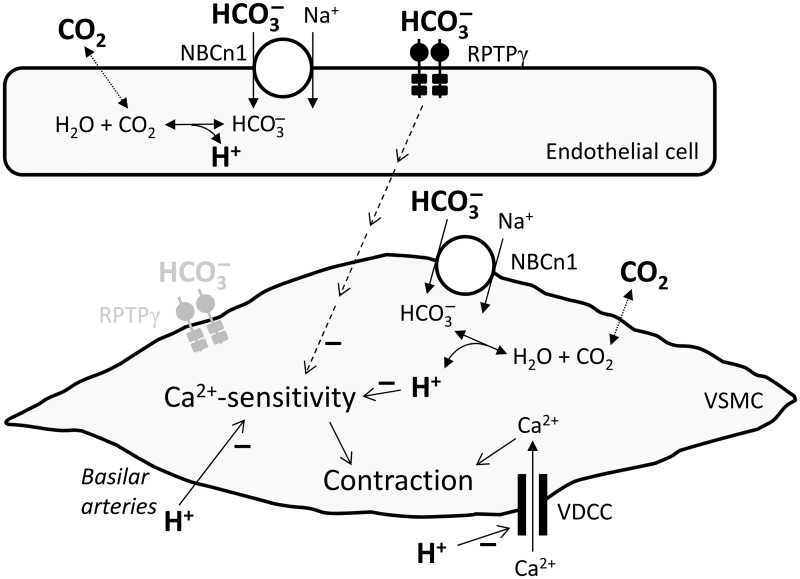
We investigate sensing and signaling mechanisms for H(+), [Formula: see text] and CO2 in basilar arteries using out-of-equilibrium solutions. Selectively varying pHo, [[Formula: see text]]o, or pCO2, we find: (a) lowering pHo attenuates vasoconstriction and vascular smooth muscle cell (VSMC) Ca(2+)-responses whereas raising pHo augments vasoconstriction independently of VSMC [Ca(2+)]i, (b) lowering [[Formula: see text]]o increases arterial agonist-sensitivity of tone development without affecting VSMC [Ca(2+)]i but c) no evidence that CO2 has direct net vasomotor effects. Receptor protein tyrosine phosphatase (RPTP)γ is transcribed in endothelial cells, and direct vasomotor effects of [Formula: see text] are absent in arteries from RPTPγ-knockout mice. At pHo 7.4, selective changes in [[Formula: see text]]o or pCO2 have little effect on pHi At pHo 7.1, decreased [[Formula: see text]]o or increased pCO2 causes intracellular acidification, which attenuates vasoconstriction. Under equilibrated conditions, anti-contractile effects of CO2/[Formula: see text] are endothelium-dependent and absent in arteries from RPTPγ-knockout mice. With CO2/[Formula: see text] present, contractile responses to agonist-stimulation are potentiated in arteries from RPTPγ-knockout compared to wild-type mice, and this difference is larger for respiratory than metabolic acidosis. In conclusion, decreased pHo and pHi inhibit vasoconstriction, whereas decreased [[Formula: see text]]o promotes vasoconstriction through RPTPγ-dependent changes in VSMC Ca(2+)-sensitivity. [Formula: see text] serves dual roles, providing substrate for pHi-regulating membrane transporters and modulating arterial responses to acid-base disturbances.
Keywords: Vascular biology; basic science; calcium imaging; confocal microscopy; electrophysiology; endothelium; experimental; pH; physiology; receptors; smooth muscle.
© The Author(s) 2015.
Figures
References
-
- Kontos HA. Regulation of the cerebral circulation. Ann Rev Physiol 1981; 43: 397–407. - PubMed
-
- Zhao J, Hogan EM, Bevensee MO, et al. Out-of-equilibrium CO2/ solutions and their use in characterizing a new K/HCO3 cotransporter. Nature 1995; 374: 636–639. - PubMed
-
- Peng HL, Ivarsen A, Nilsson H, et al. On the cellular mechanism for the effect of acidosis on vascular tone. Acta Physiol Scand 1998; 164: 517–525. - PubMed
-
- Boedtkjer E, Praetorius J, Aalkjaer C. NBCn1 (slc4a7) mediates the Na+-dependent bicarbonate transport important for regulation of intracellular pH in mouse vascular smooth muscle cells. Circ Res 2006; 98: 515–523. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
