

Gain-of-Function Mutation W493R in the Epithelial Sodium Channel Allosterically Reconfigures Intersubunit Coupling
- PMID: 26668308
- PMCID: PMC4759151
- DOI: 10.1074/jbc.M115.678052
Gain-of-Function Mutation W493R in the Epithelial Sodium Channel Allosterically Reconfigures Intersubunit Coupling
Abstract
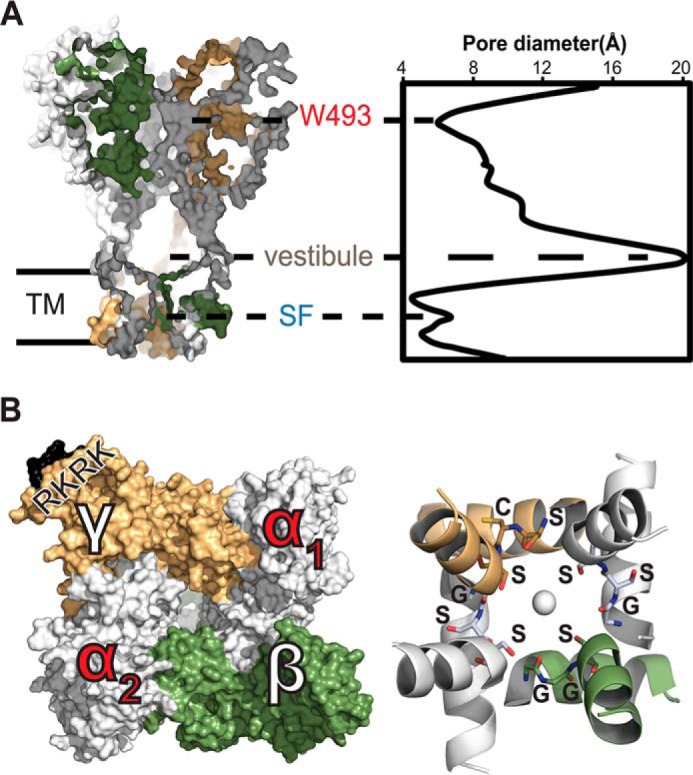
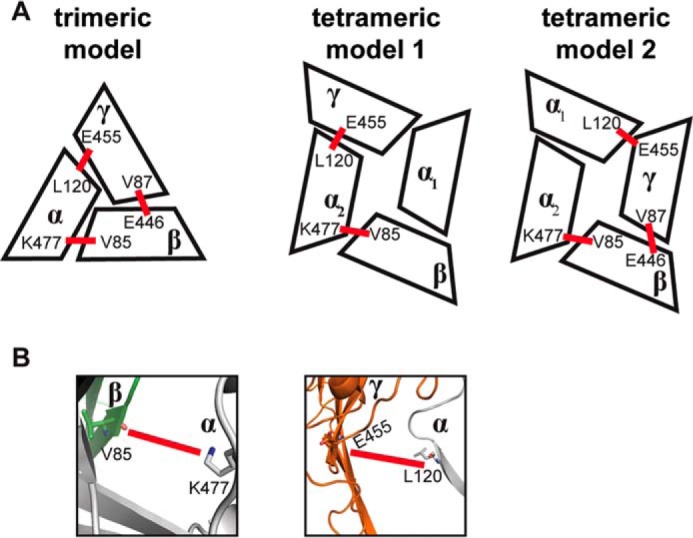
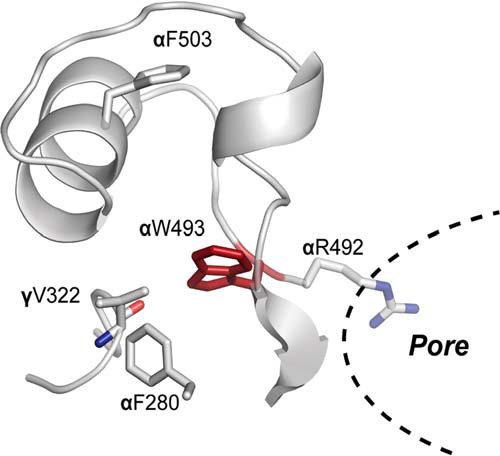
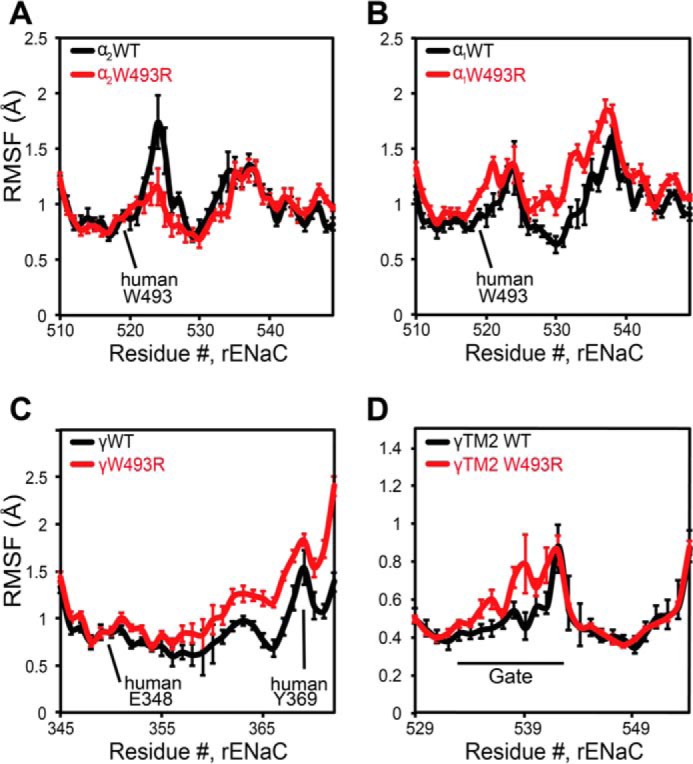
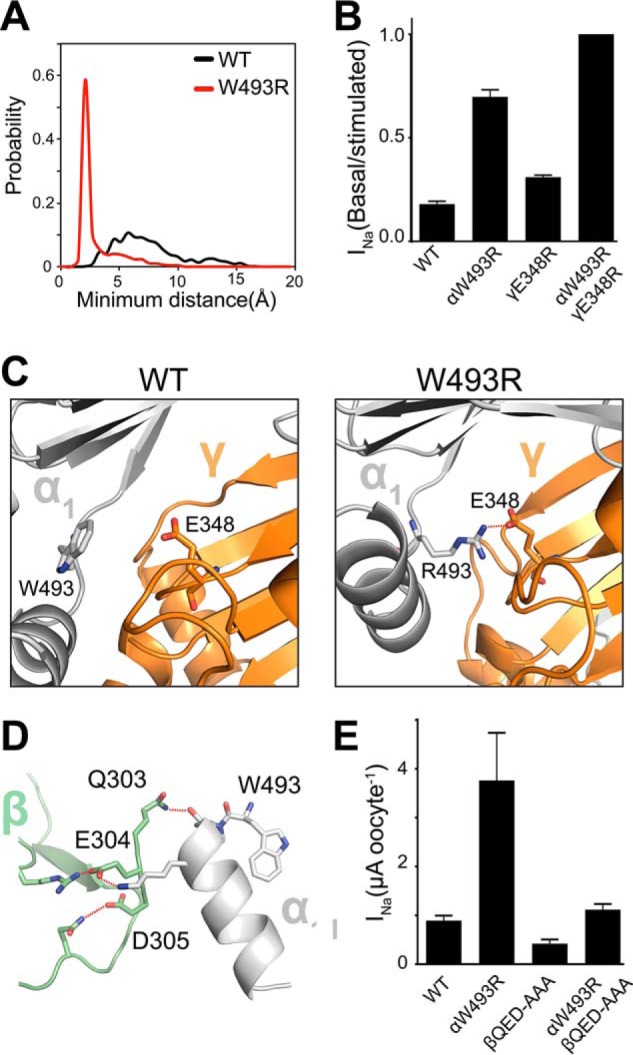
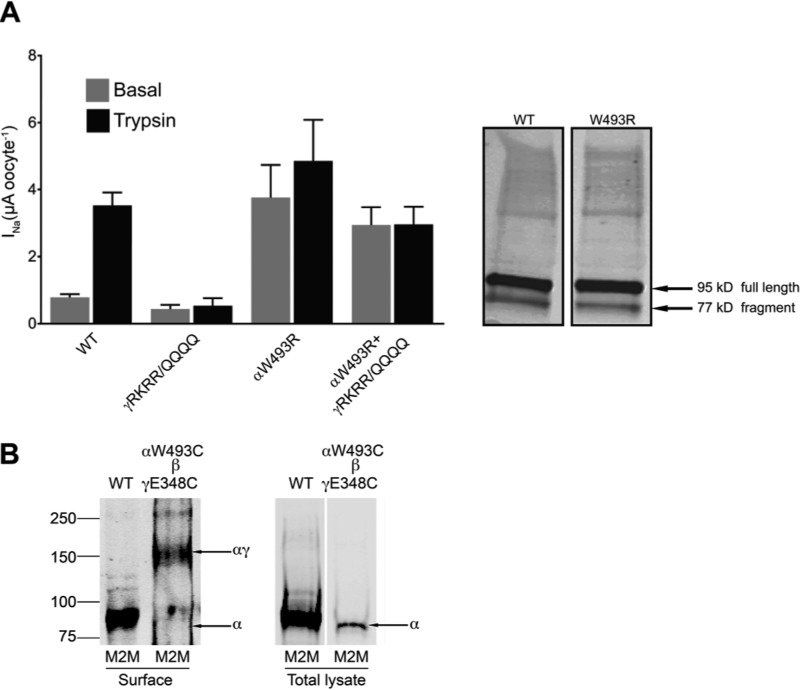
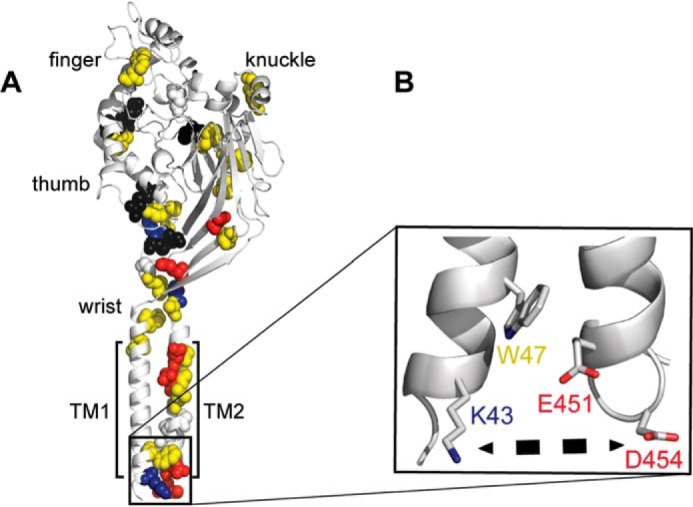
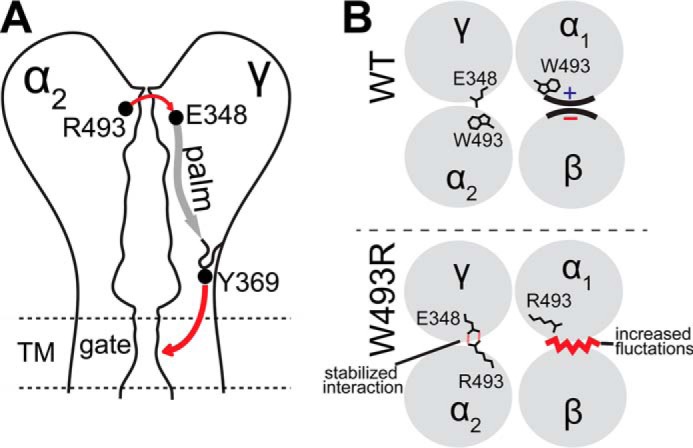
Sodium absorption in epithelial cells is rate-limited by the epithelial sodium channel (ENaC) activity in lung, kidney, and the distal colon. Pathophysiological conditions, such as cystic fibrosis and Liddle syndrome, result from water-electrolyte imbalance partly due to malfunction of ENaC regulation. Because the quaternary structure of ENaC is yet undetermined, the bases of pathologically linked mutations in ENaC subunits α, β, and γ are largely unknown. Here, we present a structural model of heterotetrameric ENaC α1βα2γ that is consistent with previous cross-linking results and site-directed mutagenesis experiments. By using this model, we show that the disease-causing mutation αW493R rewires structural dynamics of the intersubunit interfaces α1β and α2γ. Changes in dynamics can allosterically propagate to the channel gate. We demonstrate that cleavage of the γ-subunit, which is critical for full channel activation, does not mediate activation of ENaC by αW493R. Our molecular dynamics simulations led us to identify a channel-activating electrostatic interaction between α2Arg-493 and γGlu-348 at the α2γ interface. By neutralizing a sodium-binding acidic patch at the α1β interface, we reduced ENaC activation of αW493R by more than 2-fold. By combining homology modeling, molecular dynamics, cysteine cross-linking, and voltage clamp experiments, we propose a dynamics-driven model for the gain-of-function in ENaC by αW493R. Our integrated computational and experimental approach advances our understanding of structure, dynamics, and function of ENaC in its disease-causing state.
Keywords: allosteric activation; channelopathies; electrophysiology; ion channel; molecular docking; molecular dynamics; protein-protein interaction; tetramer model.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Bhalla V., and Hallows K. R. (2008) Mechanisms of ENaC regulation and clinical implications. J. Am. Soc. Nephrol. 19, 1845–1854 - PubMed
-
- Azad A. K., Rauh R., Vermeulen F., Jaspers M., Korbmacher J., Boissier B., Bassinet L., Fichou Y., des Georges M., Stanke F., De Boeck K., Dupont L., Balascáková M., Hjelte L., Lebecque P., et al. (2009) Mutations in the amiloride-sensitive epithelial sodium channel in patients with cystic fibrosis-like disease. Hum. Mutat 30, 1093–1103 - PubMed
-
- Howsham C., and Danahay H. (2014) in Ion Channel Drug Discovery (Cox B., and Gosling M., eds) pp. 135–155, Royal Society of Chemistry, London
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
