

Origin and diversification of wings: Insights from a neopteran insect
- PMID: 26668365
- PMCID: PMC4702999
- DOI: 10.1073/pnas.1509517112
Origin and diversification of wings: Insights from a neopteran insect
Abstract
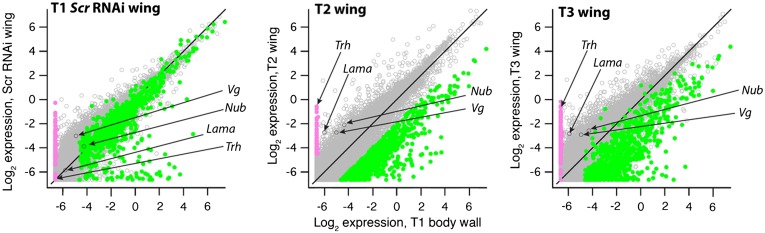
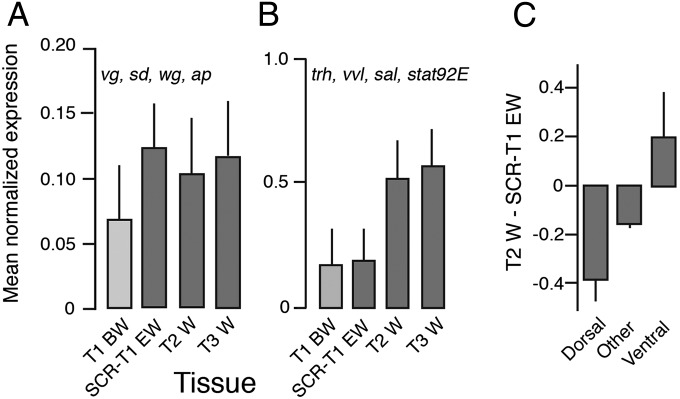
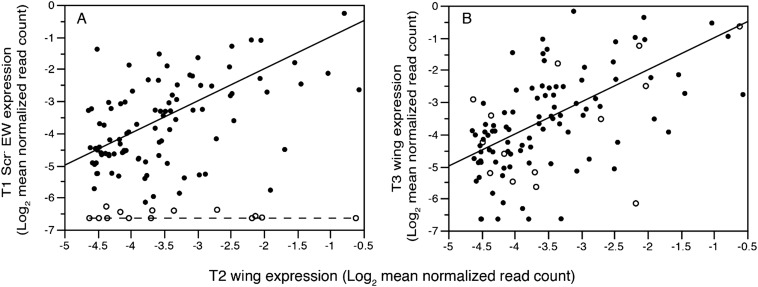
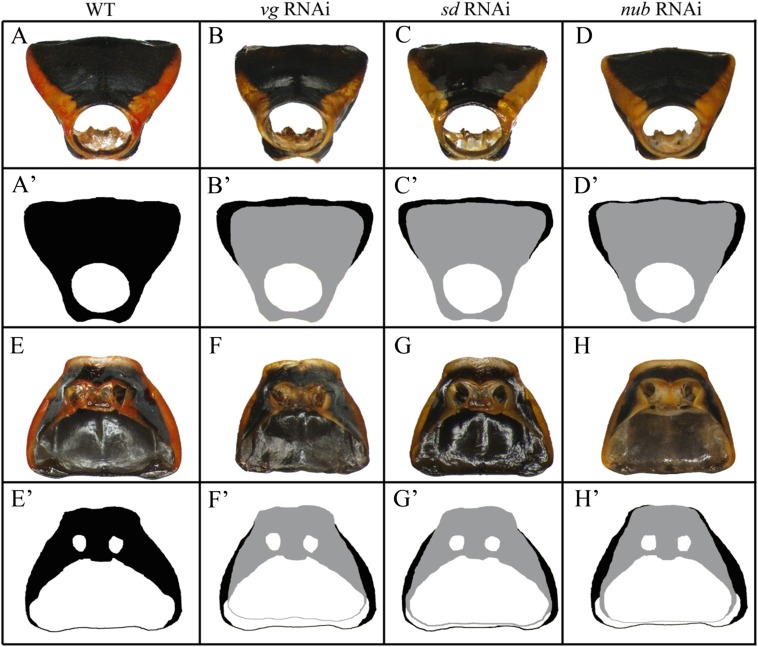
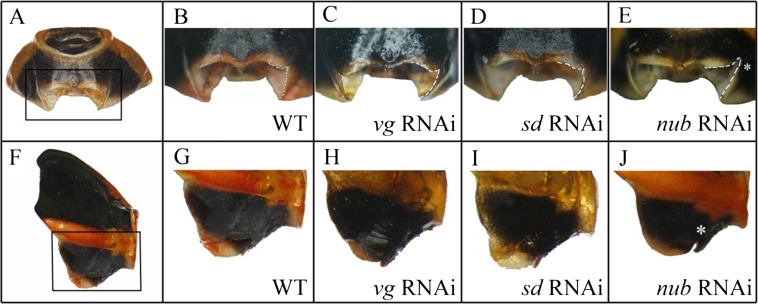
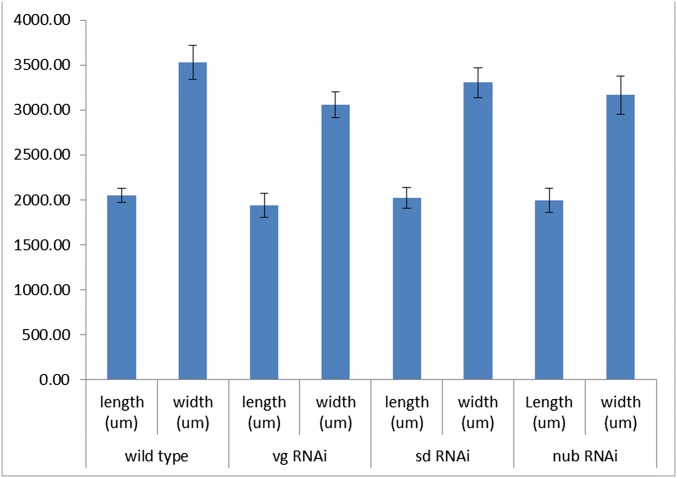
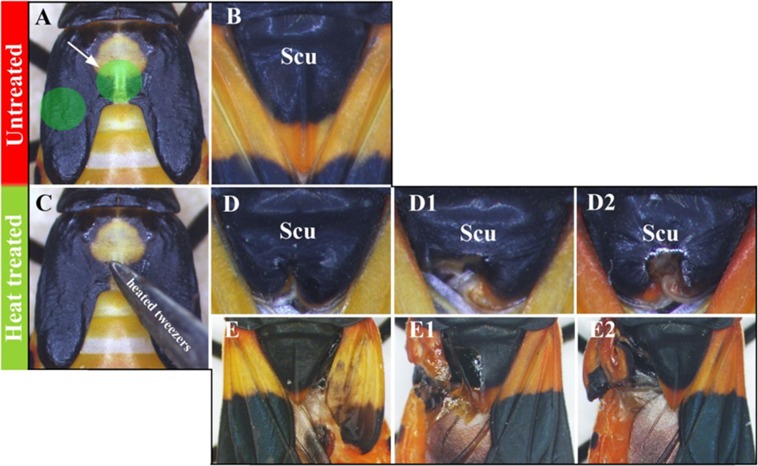
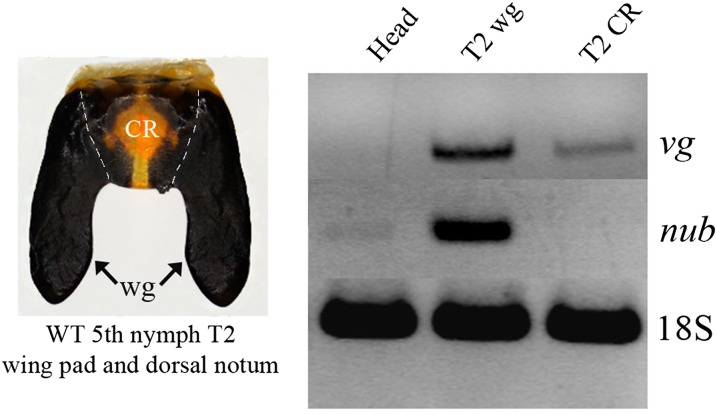
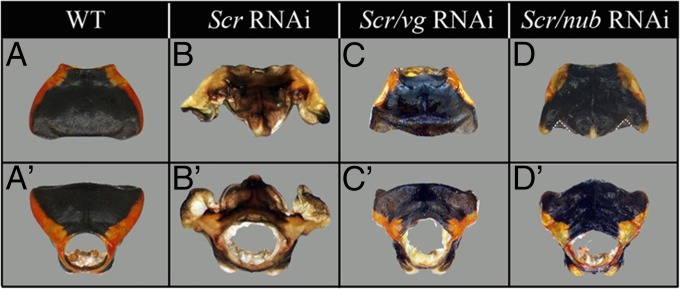
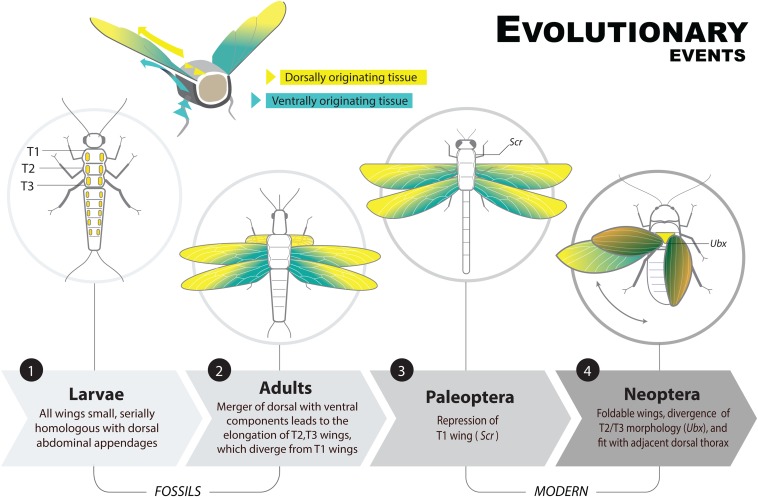
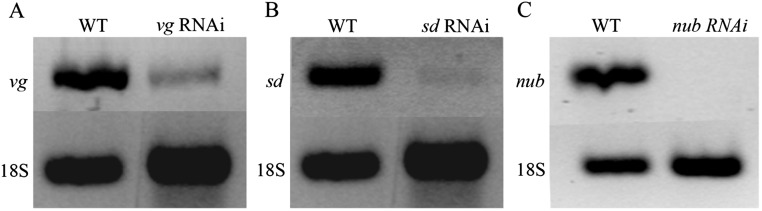
Winged insects underwent an unparalleled evolutionary radiation, but mechanisms underlying the origin and diversification of wings in basal insects are sparsely known compared with more derived holometabolous insects. In the neopteran species Oncopeltus fasciatus, we manipulated wing specification genes and used RNA-seq to obtain both functional and genomic perspectives. Combined with previous studies, our results suggest the following key steps in wing origin and diversification. First, a set of dorsally derived outgrowths evolved along a number of body segments including the first thoracic segment (T1). Homeotic genes were subsequently co-opted to suppress growth of some dorsal flaps in the thorax and abdomen. In T1 this suppression was accomplished by Sex combs reduced, that when experimentally removed, results in an ectopic T1 flap similar to prothoracic winglets present in fossil hemipteroids and other early insects. Global gene-expression differences in ectopic T1 vs. T2/T3 wings suggest that the transition from flaps to wings required ventrally originating cells, homologous with those in ancestral arthropod gill flaps/epipods, to migrate dorsally and fuse with the dorsal flap tissue thereby bringing new functional gene networks; these presumably enabled the T2/T3 wing's increased size and functionality. Third, "fused" wings became both the wing blade and surrounding regions of the dorsal thorax cuticle, providing tissue for subsequent modifications including wing folding and the fit of folded wings. Finally, Ultrabithorax was co-opted to uncouple the morphology of T2 and T3 wings and to act as a general modifier of hindwings, which in turn governed the subsequent diversification of lineage-specific wing forms.
Keywords: RNA-seq; Sex combs reduced; Ultrabithorax; vestigial; wing origins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Grimaldi D, Engel MS. Evolution of the Insects. Cambridge Univ Press; New York: 2005. Insects take to the skies; pp. 155–187.
-
- Wootton RJ, Kukalová-Peck J. Flight adaptations in Palaeozoic Palaeoptera (Insecta) Biol Rev Camb Philos Soc. 2000;75(1):129–167. - PubMed
-
- Dudley R. The Biomechanics of Insect Flight: Form, Function, and Evolution. Princeton Univ Press; Princeton, NJ: 2000.
-
- Jockusch EL, Nagy LM. Insect evolution: How did insect wings originate? Curr Biol. 1997;7(6):R358–R361. - PubMed
-
- Tomoyasu Y, Wheeler SR, Denell RE. Ultrabithorax is required for membranous wing identity in the beetle Tribolium castaneum. Nature. 2005;433(7026):643–647. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
