

AMP-activated Protein Kinase Deficiency Blocks the Hypoxic Ventilatory Response and Thus Precipitates Hypoventilation and Apnea
- PMID: 26669206
- PMCID: PMC4872655
- DOI: 10.1164/rccm.201508-1667OC
AMP-activated Protein Kinase Deficiency Blocks the Hypoxic Ventilatory Response and Thus Precipitates Hypoventilation and Apnea
Abstract
Rationale: Modulation of breathing by hypoxia accommodates variations in oxygen demand and supply during, for example, sleep and ascent to altitude, but the precise molecular mechanisms of this phenomenon remain controversial. Among the genes influenced by natural selection in high-altitude populations is one for the adenosine monophosphate-activated protein kinase (AMPK) α1-catalytic subunit, which governs cell-autonomous adaptations during metabolic stress.
Objectives: We investigated whether AMPK-α1 and/or AMPK-α2 are required for the hypoxic ventilatory response and the mechanism of ventilatory dysfunctions arising from AMPK deficiency.
Methods: We used plethysmography, electrophysiology, functional magnetic resonance imaging, and immediate early gene (c-fos) expression to assess the hypoxic ventilatory response of mice with conditional deletion of the AMPK-α1 and/or AMPK-α2 genes in catecholaminergic cells, which compose the hypoxia-responsive respiratory network from carotid body to brainstem.
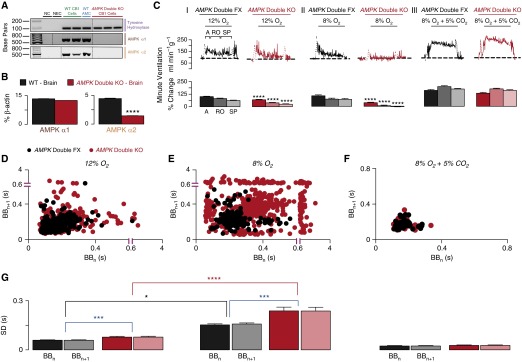
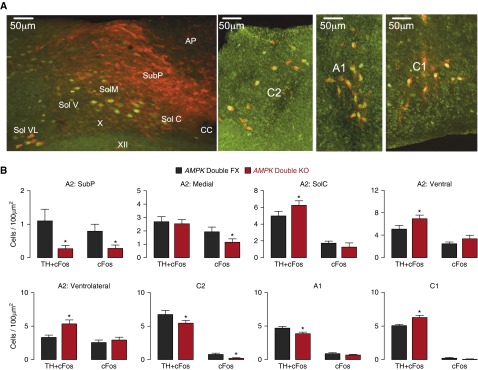
Measurements and main results: AMPK-α1 and AMPK-α2 deletion virtually abolished the hypoxic ventilatory response, and ventilatory depression during hypoxia was exacerbated under anesthesia. Rather than hyperventilating, mice lacking AMPK-α1 and AMPK-α2 exhibited hypoventilation and apnea during hypoxia, with the primary precipitant being loss of AMPK-α1 expression. However, the carotid bodies of AMPK-knockout mice remained exquisitely sensitive to hypoxia, contrary to the view that the hypoxic ventilatory response is determined solely by increased carotid body afferent input to the brainstem. Regardless, functional magnetic resonance imaging and c-fos expression revealed reduced activation by hypoxia of well-defined dorsal and ventral brainstem nuclei.
Conclusions: AMPK is required to coordinate the activation by hypoxia of brainstem respiratory networks, and deficiencies in AMPK expression precipitate hypoventilation and apnea, even when carotid body afferent input is normal.
Keywords: adenosine monophosphate–activated protein kinase; apnea; brainstem; hypoxia; ventilation.
Figures





Comment in
-
The Energy Crisis Revisited: AMP-activated Protein Kinase and the Mammalian Hypoxic Ventilatory Response.Am J Respir Crit Care Med. 2016 May 1;193(9):945-6. doi: 10.1164/rccm.201512-2323ED. Am J Respir Crit Care Med. 2016. PMID: 27128704 No abstract available.
References
-
- Woods A, Dickerson K, Heath R, Hong SP, Momcilovic M, Johnstone SR, Carlson M, Carling D. Ca2+/calmodulin-dependent protein kinase kinase-β acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab. 2005;2:21–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
