

Formal models in animal-metacognition research: the problem of interpreting animals' behavior
- PMID: 26669600
- PMCID: PMC4909597
- DOI: 10.3758/s13423-015-0985-2
Formal models in animal-metacognition research: the problem of interpreting animals' behavior
Abstract
Ongoing research explores whether animals have precursors to metacognition-that is, the capacity to monitor mental states or cognitive processes. Comparative psychologists have tested apes, monkeys, rats, pigeons, and a dolphin using perceptual, memory, foraging, and information-seeking paradigms. The consensus is that some species have a functional analog to human metacognition. Recently, though, associative modelers have used formal-mathematical models hoping to describe animals' "metacognitive" performances in associative-behaviorist ways. We evaluate these attempts to reify formal models as proof of particular explanations of animal cognition. These attempts misunderstand the content and proper application of models. They embody mistakes of scientific reasoning. They blur fundamental distinctions in understanding animal cognition. They impede theoretical development. In contrast, an energetic empirical enterprise is achieving strong success in describing the psychology underlying animals' metacognitive performances. We argue that this careful empirical work is the clear path to useful theoretical development. The issues raised here about formal modeling-in the domain of animal metacognition-potentially extend to biobehavioral research more broadly.
Keywords: Associative learning; Comparative psychology; Metacognition; Metamemory; Modeling; Primate cognition.
Figures
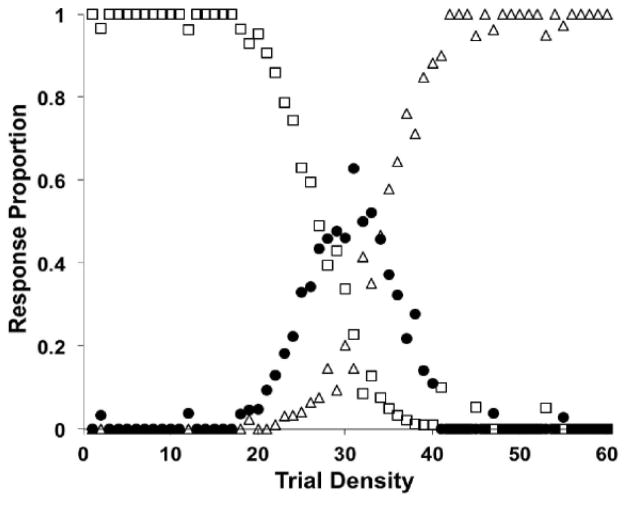
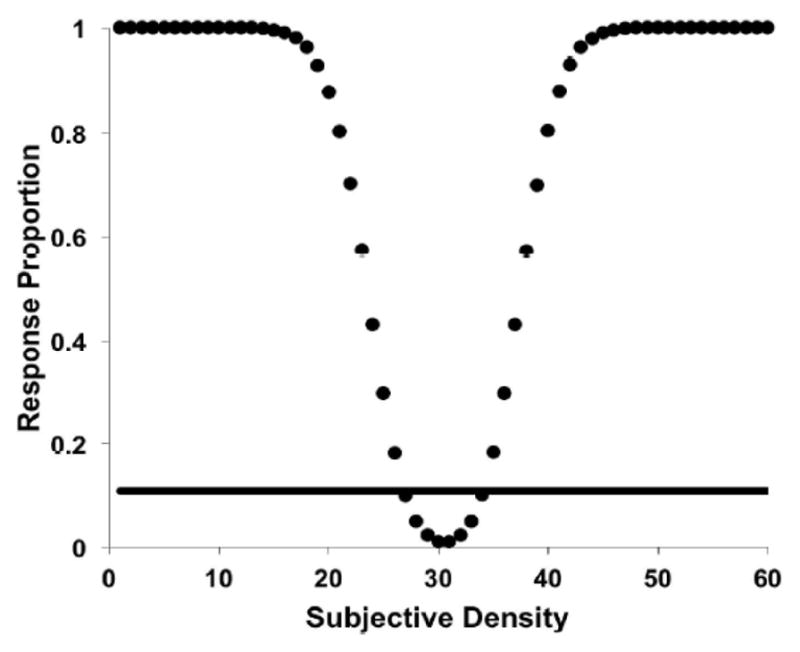
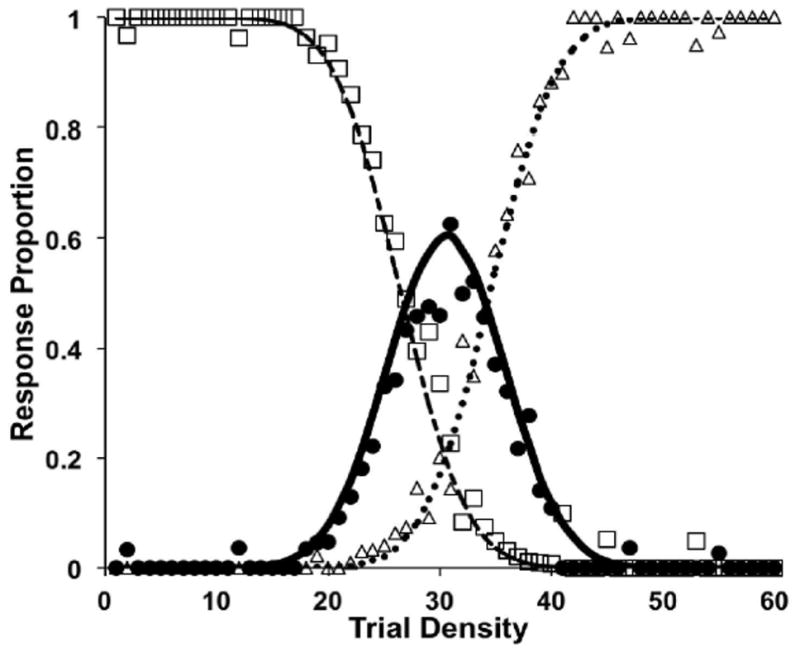
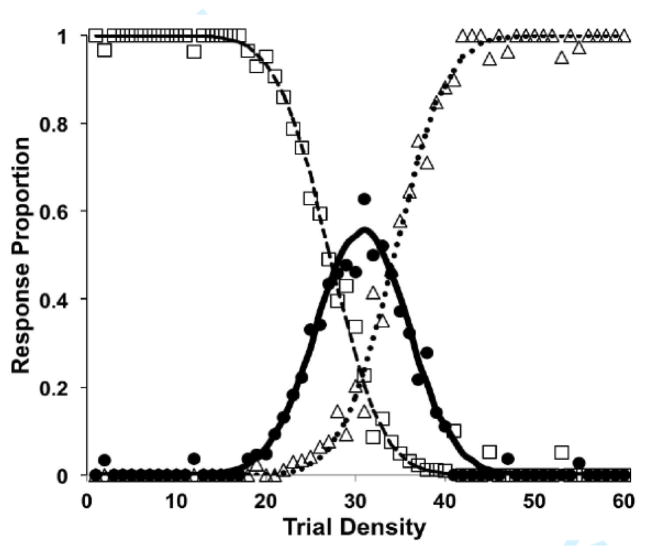
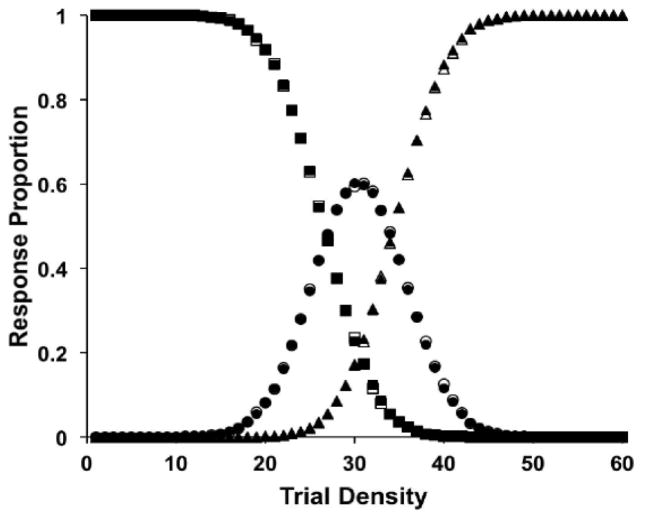
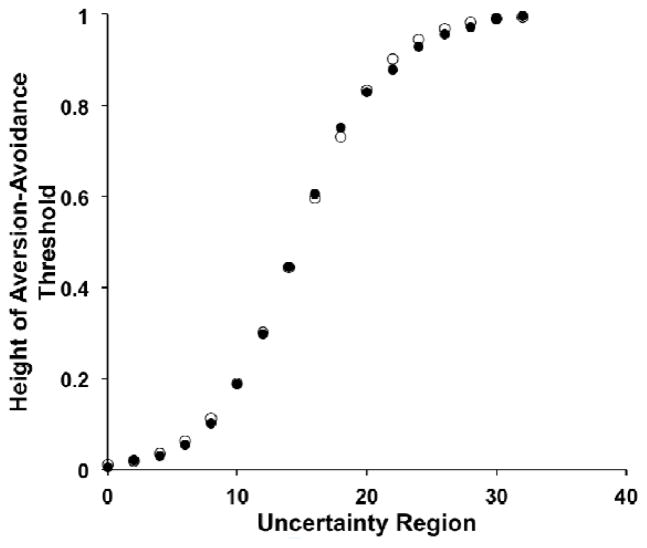
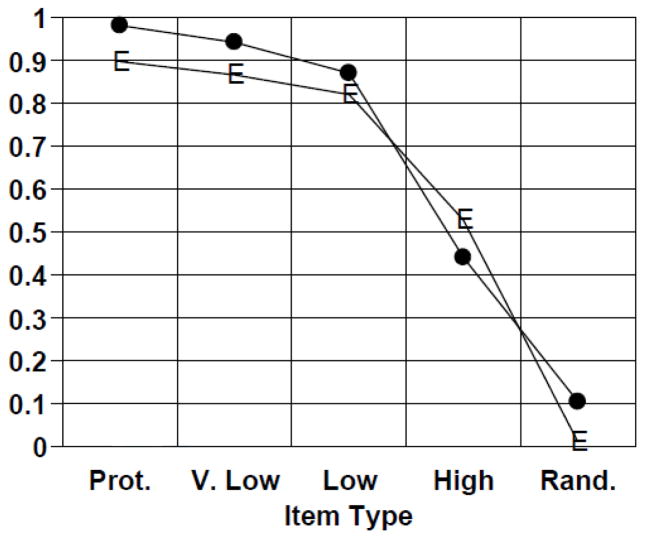
Similar articles
-
Animal metacognition: a tale of two comparative psychologies.J Comp Psychol. 2014 May;128(2):115-31. doi: 10.1037/a0033105. Epub 2013 Aug 19. J Comp Psychol. 2014. PMID: 23957740 Free PMC article. Review.
-
The comparative psychology of uncertainty monitoring and metacognition.Behav Brain Sci. 2003 Jun;26(3):317-39; discussion 340-73. doi: 10.1017/s0140525x03000086. Behav Brain Sci. 2003. PMID: 14968691
-
The study of animal metacognition.Trends Cogn Sci. 2009 Sep;13(9):389-96. doi: 10.1016/j.tics.2009.06.009. Epub 2009 Aug 31. Trends Cogn Sci. 2009. PMID: 19726218 Review.
-
The highs and lows of theoretical interpretation in animal-metacognition research.Philos Trans R Soc Lond B Biol Sci. 2012 May 19;367(1594):1297-309. doi: 10.1098/rstb.2011.0366. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 22492748 Free PMC article. Review.
-
Explicit memory and cognition in monkeys.Neuropsychologia. 2020 Feb 17;138:107326. doi: 10.1016/j.neuropsychologia.2019.107326. Epub 2020 Jan 7. Neuropsychologia. 2020. PMID: 31917205 Free PMC article. Review.
Cited by
-
Are There Differences in "Intelligence" Between Nonhuman Species? The Role of Contextual Variables.Front Psychol. 2020 Aug 21;11:2072. doi: 10.3389/fpsyg.2020.02072. eCollection 2020. Front Psychol. 2020. PMID: 32973624 Free PMC article. Review.
-
Computational animal welfare: towards cognitive architecture models of animal sentience, emotion and wellbeing.R Soc Open Sci. 2020 Dec 23;7(12):201886. doi: 10.1098/rsos.201886. eCollection 2020 Dec. R Soc Open Sci. 2020. PMID: 33489298 Free PMC article. Review.
-
A metacognitive illusion in monkeys.Proc Biol Sci. 2017 Sep 13;284(1862):20171541. doi: 10.1098/rspb.2017.1541. Proc Biol Sci. 2017. PMID: 28878068 Free PMC article.
-
Metacognition in dogs: Do dogs know they could be wrong?Learn Behav. 2018 Dec;46(4):398-413. doi: 10.3758/s13420-018-0367-5. Learn Behav. 2018. PMID: 30421122 Free PMC article.
-
Testing for Metacognitive Responding Using an Odor-based Delayed Match-to-Sample Test in Rats.J Vis Exp. 2018 Jun 18;(136):57489. doi: 10.3791/57489. J Vis Exp. 2018. PMID: 29985304 Free PMC article.
References
-
- Arbuthnott GW, Ingham CA, Wickens JR. Dopamine and synaptic plasticity in the neostriatum. Journal of Anatomy. 2000;196:587–596. http://dx.doi.org/10.1046/j.1469-7580.2000.19640587.x. - DOI - PMC - PubMed
-
- Balcomb FK, Gerken L. Three-year-old children can access their own memory to guide responses on a visual matching task. Developmental Science. 2008;11:750–760. http://dx.doi.org/10.1111/j.1467-7687.2008.00725.x. - DOI - PubMed
-
- Basile BM, Hampton RR. Metacognition as discrimination: commentary on Smith et al. (2014) Journal of Comparative Psychology. 2014;128:135–137. http://dx.doi.org/10.1037/a0034412. - DOI - PMC - PubMed
-
- Basile BM, Hampton RR, Suomi SJ, Murray EA. An assessment of memory awareness in tufted capuchin monkeys (Cebus apella) Animal Cognition. 2009;12:169–180. http://dx.doi.org/10.1007/s10071-008-0180-1. - DOI - PMC - PubMed
-
- Basile BM, Schroeder GR, Brown EK, Templer VL, Hampton RR. Evaluation of seven hypotheses for metamemory performance in rhesus monkeys. Journal of Experimental Psychology: General. 2015;144:85–102. http://psycnet.apa.org/doi/10.1037/xge0000031. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
