

Ternary structure reveals mechanism of a membrane diacylglycerol kinase
- PMID: 26673816
- PMCID: PMC4703834
- DOI: 10.1038/ncomms10140
Ternary structure reveals mechanism of a membrane diacylglycerol kinase
Abstract
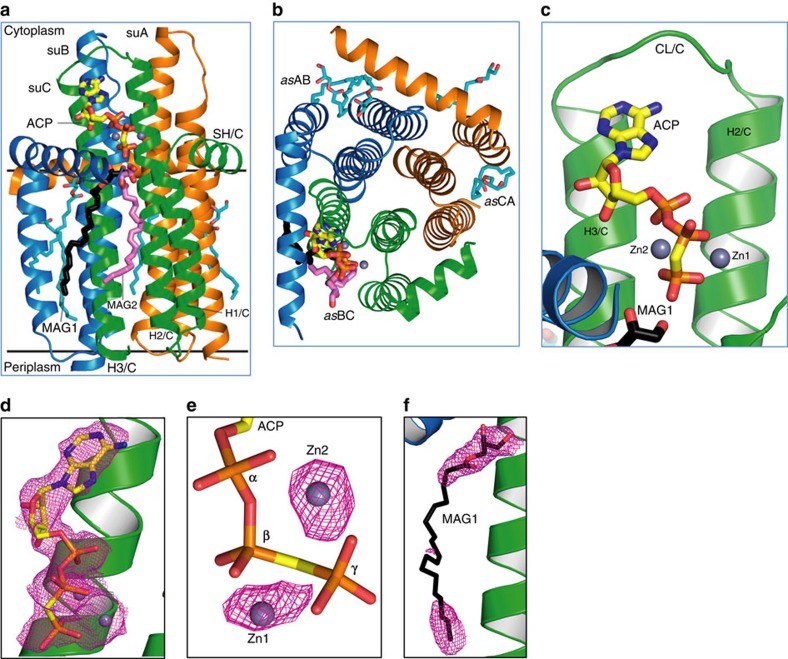
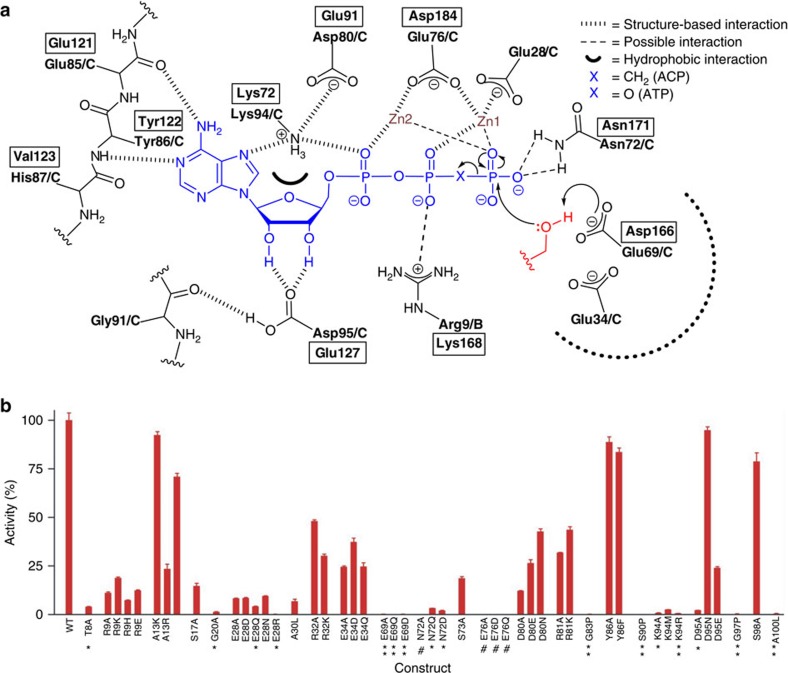
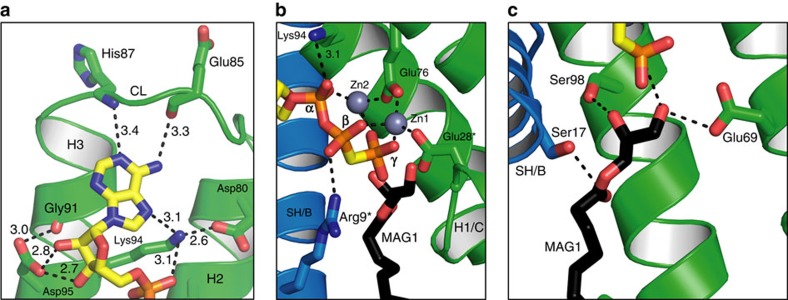
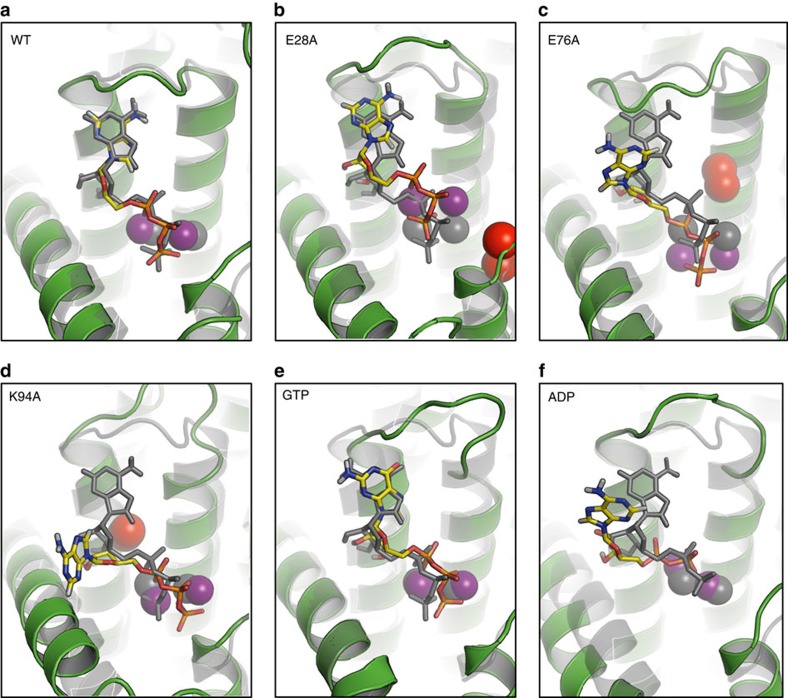
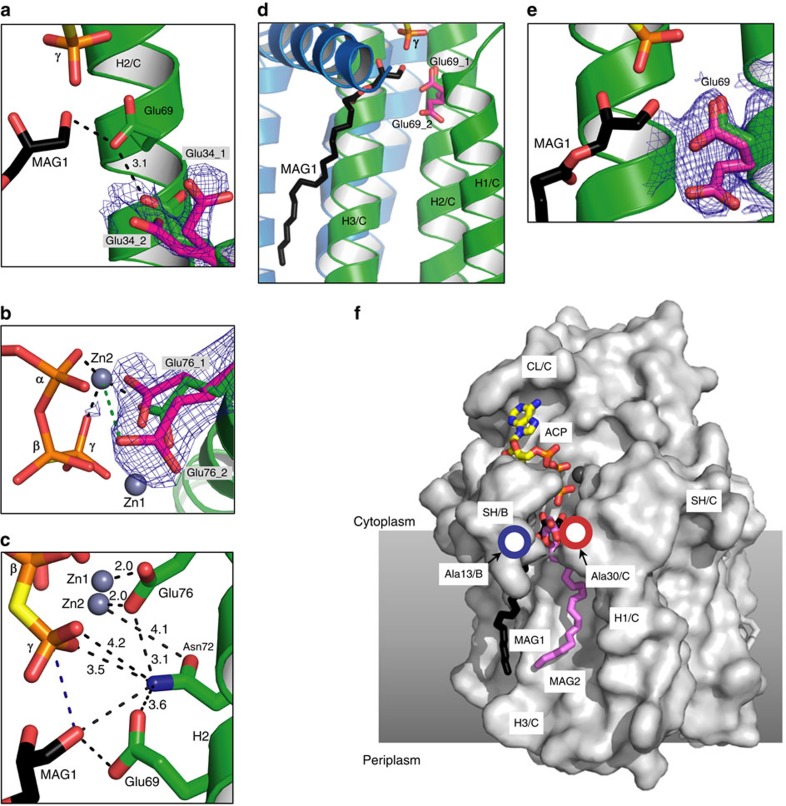
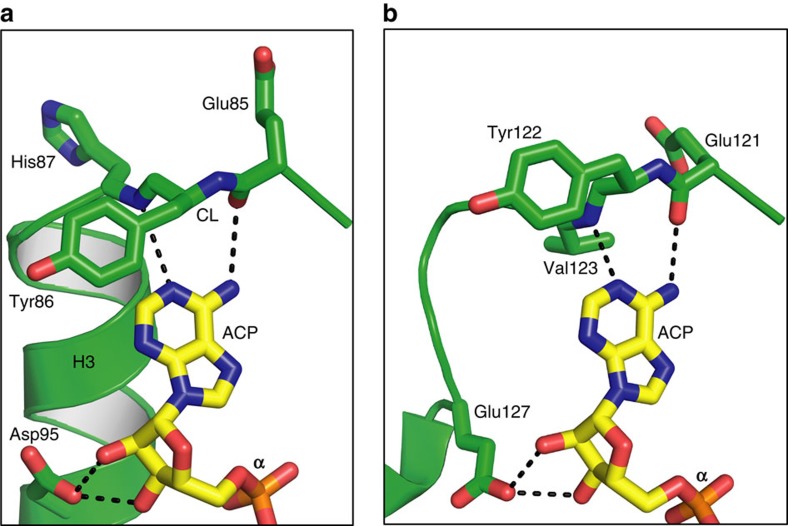
Diacylglycerol kinase catalyses the ATP-dependent conversion of diacylglycerol to phosphatidic acid in the plasma membrane of Escherichia coli. The small size of this integral membrane trimer, which has 121 residues per subunit, means that available protein must be used economically to craft three catalytic and substrate-binding sites centred about the membrane/cytosol interface. How nature has accomplished this extraordinary feat is revealed here in a crystal structure of the kinase captured as a ternary complex with bound lipid substrate and an ATP analogue. Residues, identified as essential for activity by mutagenesis, decorate the active site and are rationalized by the ternary structure. The γ-phosphate of the ATP analogue is positioned for direct transfer to the primary hydroxyl of the lipid whose acyl chain is in the membrane. A catalytic mechanism for this unique enzyme is proposed. The active site architecture shows clear evidence of having arisen by convergent evolution.
Figures
Similar articles
-
Crystal structure of the integral membrane diacylglycerol kinase.Nature. 2013 May 23;497(7450):521-4. doi: 10.1038/nature12179. Epub 2013 May 15. Nature. 2013. PMID: 23676677 Free PMC article.
-
Solution nuclear magnetic resonance structure of membrane-integral diacylglycerol kinase.Science. 2009 Jun 26;324(5935):1726-9. doi: 10.1126/science.1171716. Science. 2009. PMID: 19556511 Free PMC article.
-
Phosphoryl transfer reaction catalyzed by membrane diacylglycerol kinase: a theoretical mechanism study.Phys Chem Chem Phys. 2015 Oct 14;17(38):25228-34. doi: 10.1039/c5cp03342j. Phys Chem Chem Phys. 2015. PMID: 26352441
-
Prokaryotic diacylglycerol kinase and undecaprenol kinase.Annu Rev Biophys. 2012;41:81-101. doi: 10.1146/annurev-biophys-050511-102330. Epub 2011 Dec 20. Annu Rev Biophys. 2012. PMID: 22224599 Free PMC article. Review.
-
Molecular properties of diacylglycerol kinase-epsilon in relation to function.Chem Phys Lipids. 2015 Nov;192:100-108. doi: 10.1016/j.chemphyslip.2015.06.003. Epub 2015 Jun 29. Chem Phys Lipids. 2015. PMID: 26134136 Review.
Cited by
-
Structure Determination from Lipidic Cubic Phase Embedded Microcrystals by MicroED.Structure. 2020 Oct 6;28(10):1149-1159.e4. doi: 10.1016/j.str.2020.07.006. Epub 2020 Jul 30. Structure. 2020. PMID: 32735770 Free PMC article.
-
Multiscale Simulations of Biological Membranes: The Challenge To Understand Biological Phenomena in a Living Substance.Chem Rev. 2019 May 8;119(9):5607-5774. doi: 10.1021/acs.chemrev.8b00538. Epub 2019 Mar 12. Chem Rev. 2019. PMID: 30859819 Free PMC article.
-
Crystal structure of CmABCB1 multi-drug exporter in lipidic mesophase revealed by LCP-SFX.IUCrJ. 2021 Dec 23;9(Pt 1):134-145. doi: 10.1107/S2052252521011611. eCollection 2022 Jan 1. IUCrJ. 2021. PMID: 35059217 Free PMC article.
-
Comparison of NMR and crystal structures of membrane proteins and computational refinement to improve model quality.Proteins. 2018 Jan;86(1):57-74. doi: 10.1002/prot.25402. Epub 2017 Nov 8. Proteins. 2018. PMID: 29044728 Free PMC article.
-
Serial femtosecond crystallography.Nat Rev Methods Primers. 2022 Aug 4;2:59. doi: 10.1038/s43586-022-00141-7. Nat Rev Methods Primers. 2022. PMID: 36643971 Free PMC article.
References
-
- Badola P. & Sanders C. R. Escherichia coli diacylglycerol kinase is an evolutionarily optimized membrane enzyme and catalyzes direct phosphoryl transfer. J. Biol. Chem. 272, 24176–24182 (1997). - PubMed
-
- Lau F. W., Chen X. & Bowie J. U. Active sites of diacylglycerol kinase from Escherichia coli are shared between subunits. Biochemistry 38, 5521–5527 (1999). - PubMed
-
- Mi D., Kim H. J., Hadziselimovic A. & Sanders C. R. Irreversible misfolding of diacylglycerol kinase is independent of aggregation and occurs prior to trimerization and membrane association. Biochemistry 45, 10072–10084 (2006). - PubMed
-
- Sanders C. R. et al.. Escherichia coli diacylglycerol kinase is an alpha-helical polytopic membrane protein and can spontaneously insert into preformed lipid vesicles. Biochemistry 35, 8610–8618 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
