

Deletion of Murid Herpesvirus 4 ORF63 Affects the Trafficking of Incoming Capsids toward the Nucleus
- PMID: 26676769
- PMCID: PMC4810702
- DOI: 10.1128/JVI.02942-15
Deletion of Murid Herpesvirus 4 ORF63 Affects the Trafficking of Incoming Capsids toward the Nucleus
Abstract
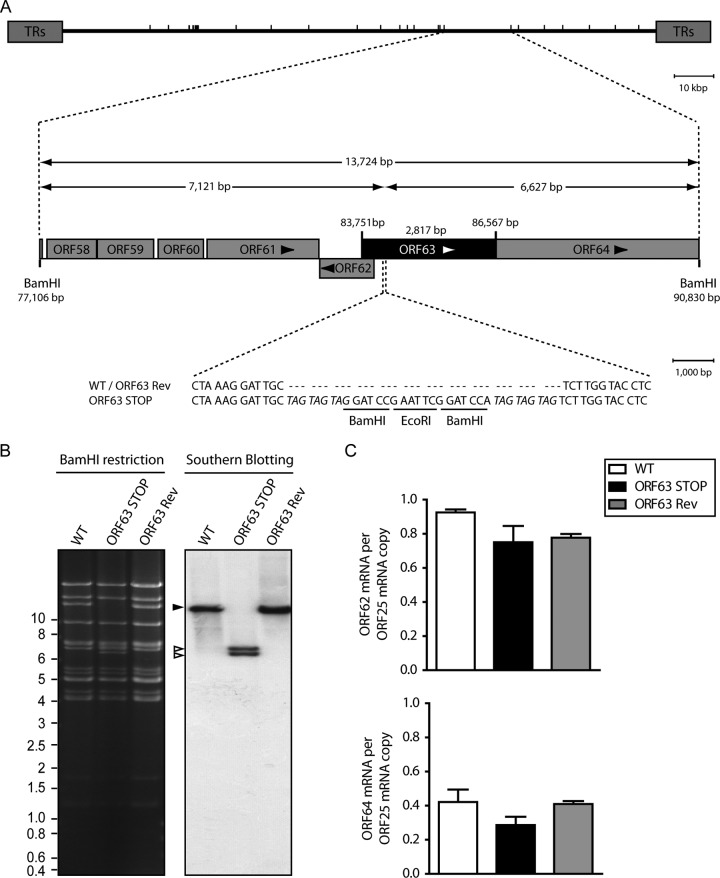
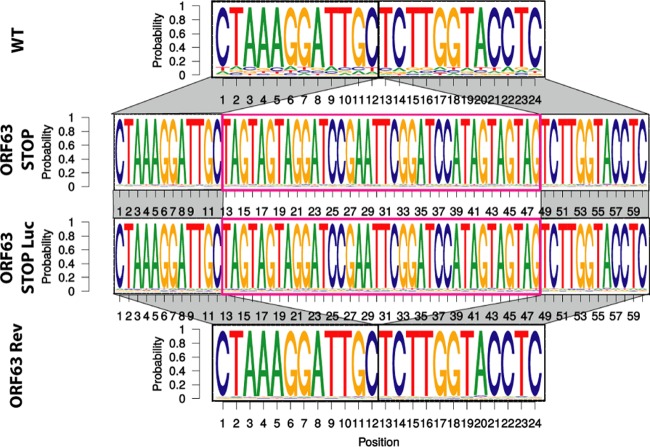
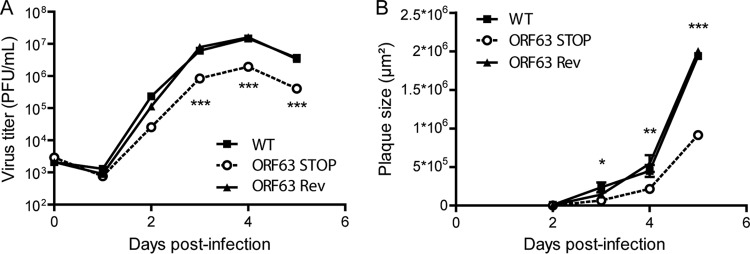
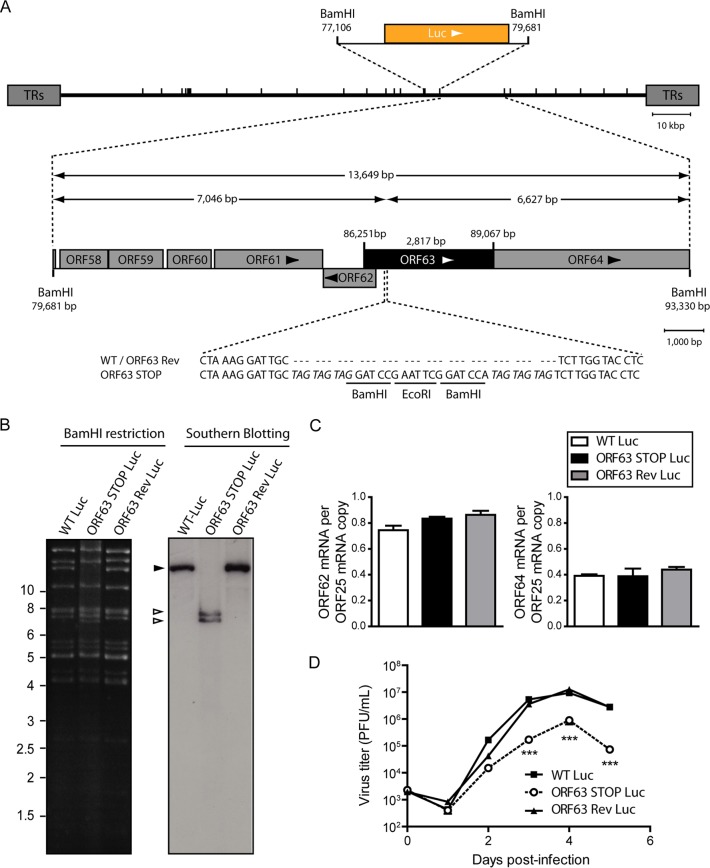
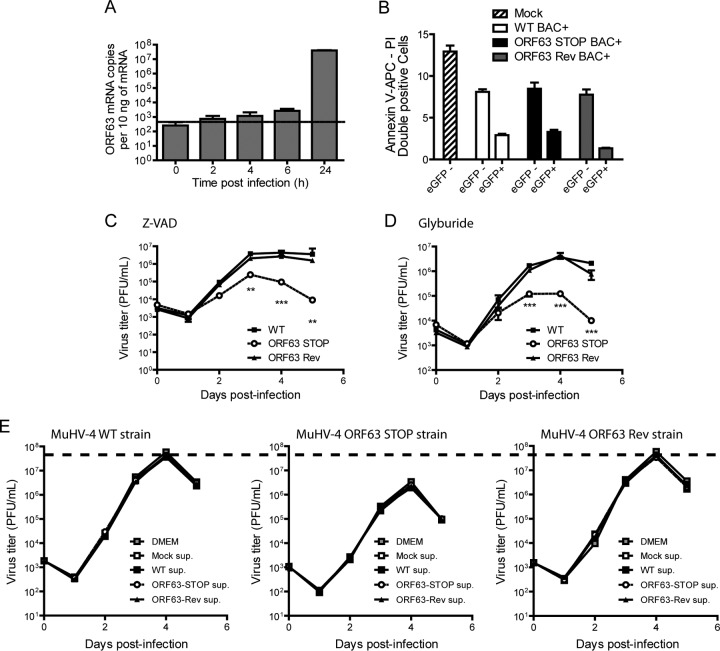
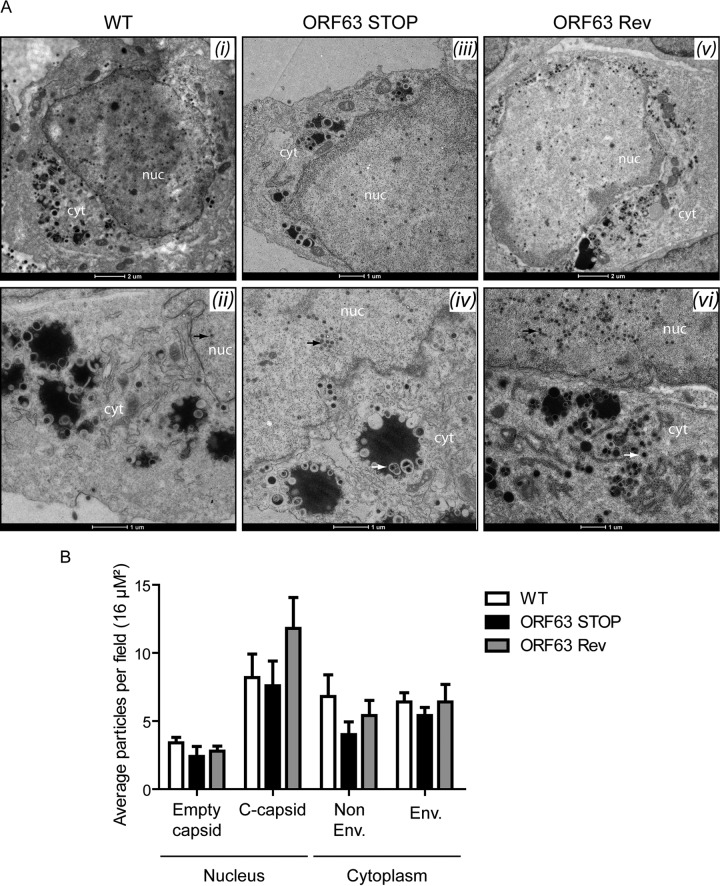
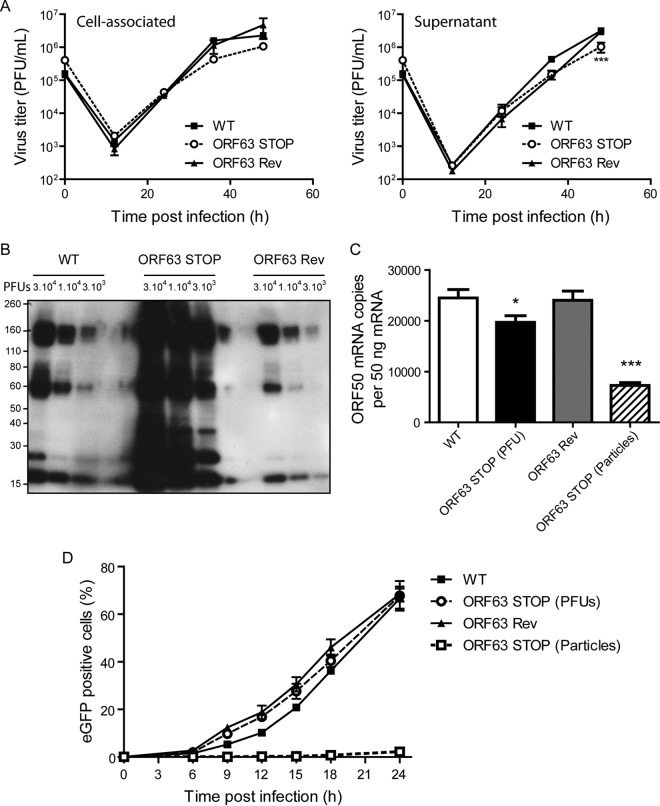
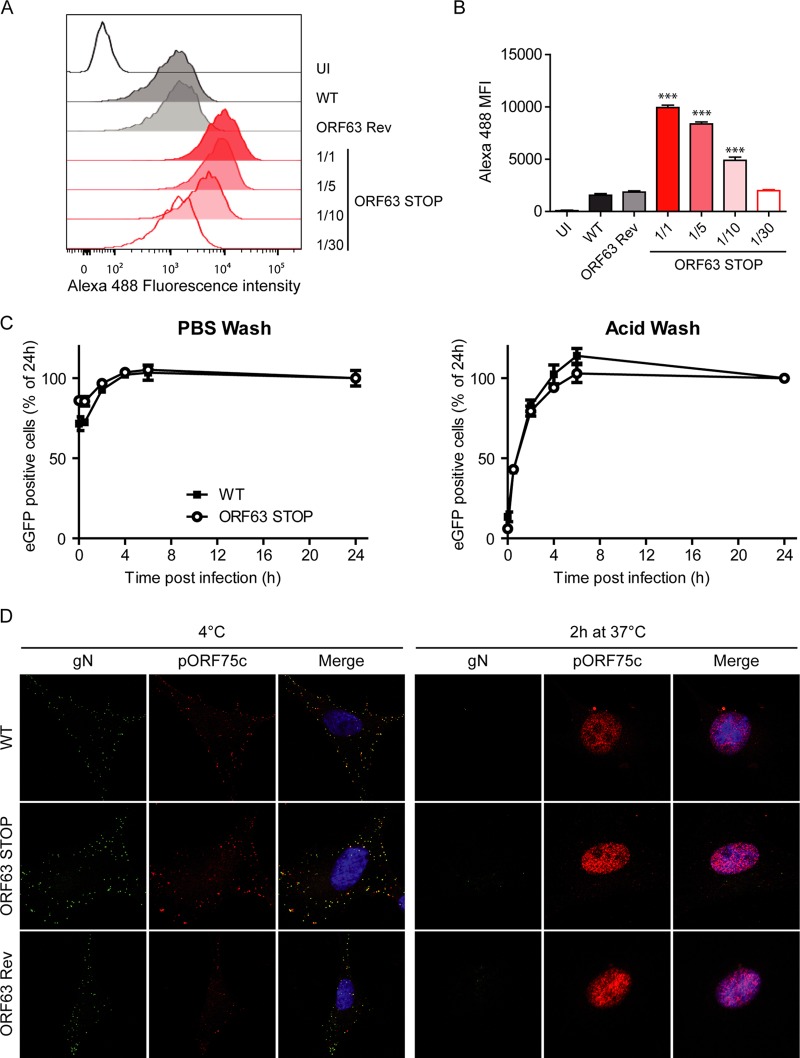
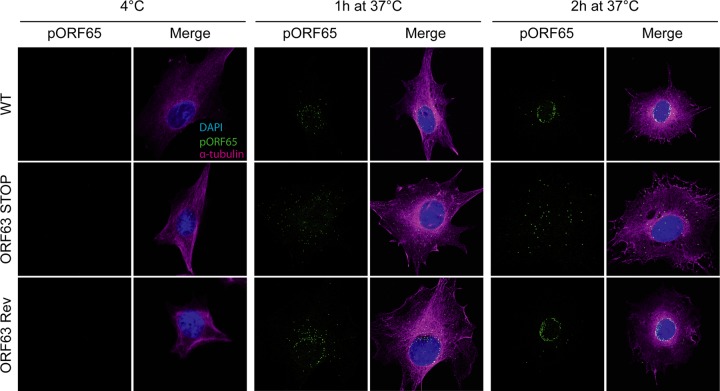
Gammaherpesviruses are important human and animal pathogens. Despite the fact that they display the classical architecture of herpesviruses, the function of most of their structural proteins is still poorly defined. This is especially true for tegument proteins. Interestingly, a potential role in immune evasion has recently been proposed for the tegument protein encoded by Kaposi's sarcoma-associated herpesvirus open reading frame 63 (ORF63). To gain insight about the roles of ORF63 in the life cycle of a gammaherpesvirus, we generated null mutations in the ORF63 gene of murid herpesvirus 4 (MuHV-4). We showed that disruption of ORF63 was associated with a severe MuHV-4 growth deficit both in vitro and in vivo. The latter deficit was mainly associated with a defect of replication in the lung but did not affect the establishment of latency in the spleen. From a functional point of view, inhibition of caspase-1 or the inflammasome did not restore the growth of the ORF63-deficient mutant, suggesting that the observed deficit was not associated with the immune evasion mechanism identified previously. Moreover, this growth deficit was also not associated with a defect in virion egress from the infected cells. In contrast, it appeared that MuHV-4 ORF63-deficient mutants failed to address most of their capsids to the nucleus during entry into the host cell, suggesting that ORF63 plays a role in capsid movement. In the future, ORF63 could therefore be considered a target to block gammaherpesvirus infection at a very early stage of the infection.
Importance: The important diseases caused by gammaherpesviruses in human and animal populations justify a better understanding of their life cycle. In particular, the role of most of their tegument proteins is still largely unknown. In this study, we used murid herpesvirus 4, a gammaherpesvirus infecting mice, to decipher the role of the protein encoded by the viral ORF63 gene. We showed that the absence of this protein is associated with a severe growth deficit both in vitro and in vivo that was mainly due to impaired migration of viral capsids toward the nucleus during entry. Together, our results provide new insights about the life cycle of gammaherpesviruses and could allow the development of new antiviral strategies aimed at blocking gammaherpesvirus infection at the very early stages.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures


References
-
- Henle G, Henle W, Clifford P, Diehl V, Kafuko GW, Kirya BG, Klein G, Morrow RH, Munube GM, Pike P, Tukei PM, Ziegler JL. 1969. Antibodies to Epstein-Barr virus in Burkitt's lymphoma and control groups. J Natl Cancer Inst 43:1147–1157. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
