

Deletion of Tricellulin Causes Progressive Hearing Loss Associated with Degeneration of Cochlear Hair Cells
- PMID: 26677943
- PMCID: PMC4683410
- DOI: 10.1038/srep18402
Deletion of Tricellulin Causes Progressive Hearing Loss Associated with Degeneration of Cochlear Hair Cells
Abstract
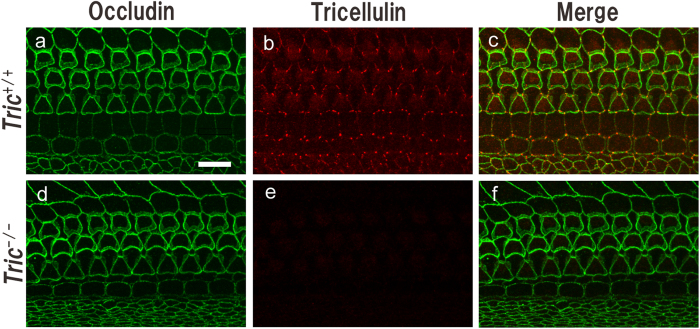
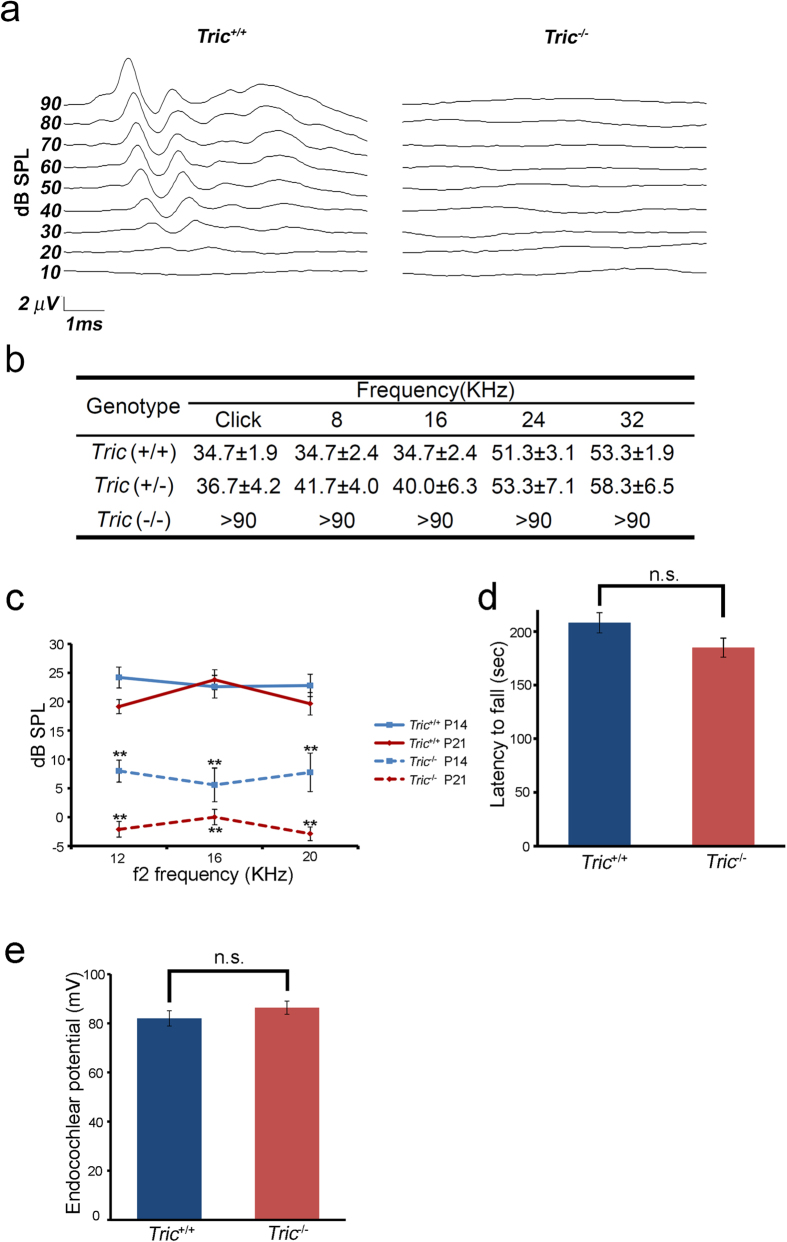
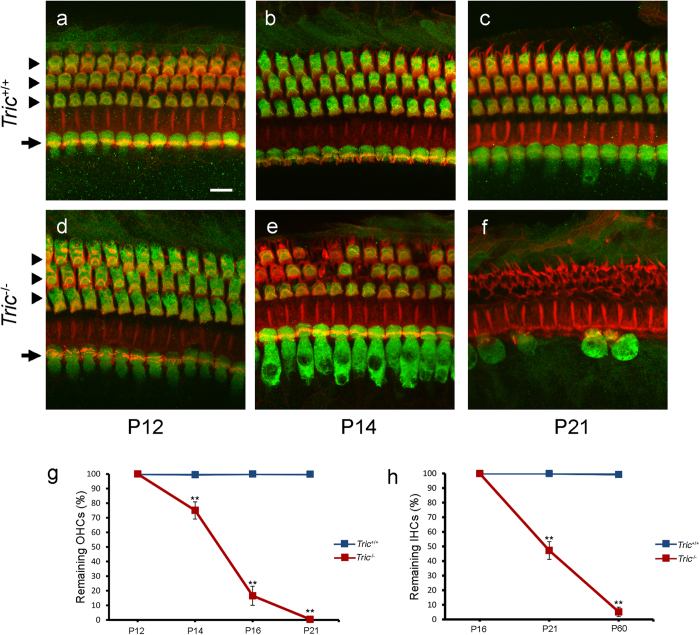
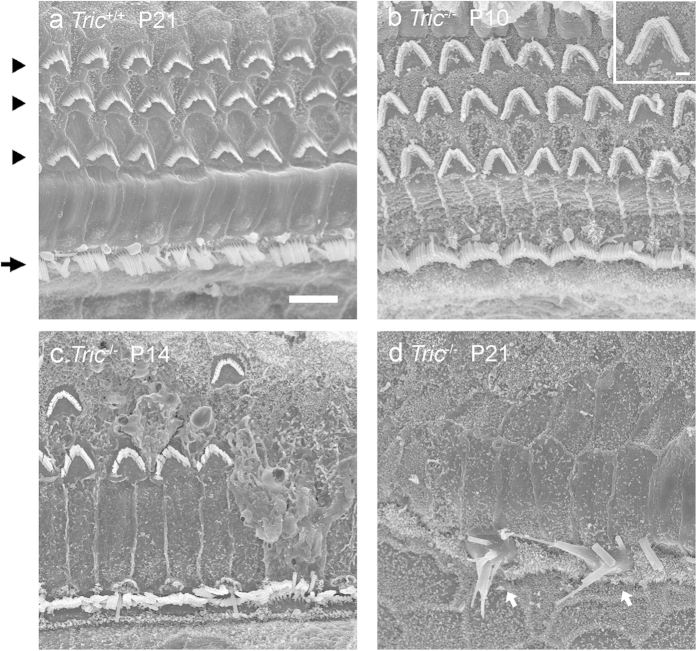
Tricellulin (also known as MARVELD2) is considered as a central component of tricellular tight junctions and is distributed among various epithelial tissues. Although mutations in the gene encoding tricellulin are known to cause deafness in humans (DFNB49) and mice, the influence of its systemic deletion in vivo remains unknown. When we generated tricellulin-knockout mice (Tric(-/-)), we found an early-onset rapidly progressive hearing loss associated with the degeneration of hair cells (HCs); however, their body size and overall appearance were normal. Tric(-/-) mice did not show any morphological change pertaining to other organs such as the gastrointestinal tract, liver, kidney, thyroid gland and heart. The endocochlear potential (EP) was normal in Tric(-/-) mice, suggesting that the tight junction barrier is maintained in the stria vascularis, where EP is generated. The degeneration of HCs, which occurred after the maturation of EP, was prevented in the culture medium with an ion concentration similar to that of the perilymph. These data demonstrate the specific requirement of tricellulin for maintaining ion homeostasis around cochlear HCs to ensure their survival. The Tric(-/-) mouse provides a new model for understanding the distinct roles of tricellulin in different epithelial systems as well as in the pathogenesis of DFNB49.
Figures
Similar articles
-
Tricellular Tight Junctions in the Inner Ear.Biomed Res Int. 2016;2016:6137541. doi: 10.1155/2016/6137541. Epub 2016 Apr 18. Biomed Res Int. 2016. PMID: 27195292 Free PMC article. Review.
-
Tricellulin deficiency affects tight junction architecture and cochlear hair cells.J Clin Invest. 2013 Sep;123(9):4036-49. doi: 10.1172/JCI69031. Epub 2013 Aug 27. J Clin Invest. 2013. PMID: 23979167 Free PMC article.
-
Deficiency of angulin-2/ILDR1, a tricellular tight junction-associated membrane protein, causes deafness with cochlear hair cell degeneration in mice.PLoS One. 2015 Mar 30;10(3):e0120674. doi: 10.1371/journal.pone.0120674. eCollection 2015. PLoS One. 2015. PMID: 25822906 Free PMC article.
-
ILDR1 null mice, a model of human deafness DFNB42, show structural aberrations of tricellular tight junctions and degeneration of auditory hair cells.Hum Mol Genet. 2015 Feb 1;24(3):609-24. doi: 10.1093/hmg/ddu474. Epub 2014 Sep 12. Hum Mol Genet. 2015. PMID: 25217574 Free PMC article.
-
A look at tricellulin and its role in tight junction formation and maintenance.Eur J Cell Biol. 2011 Oct;90(10):787-96. doi: 10.1016/j.ejcb.2011.06.005. Epub 2011 Aug 24. Eur J Cell Biol. 2011. PMID: 21868126 Review.
Cited by
-
Tricellular Tight Junctions in the Inner Ear.Biomed Res Int. 2016;2016:6137541. doi: 10.1155/2016/6137541. Epub 2016 Apr 18. Biomed Res Int. 2016. PMID: 27195292 Free PMC article. Review.
-
Angulin-1 seals tricellular contacts independently of tricellulin and claudins.J Cell Biol. 2021 Sep 6;220(9):e202005062. doi: 10.1083/jcb.202005062. Epub 2021 Jul 16. J Cell Biol. 2021. PMID: 34269802 Free PMC article.
-
Bi-Allelic MARVELD2 Variant Identified with Exome Sequencing in a Consanguineous Multiplex Ghanaian Family Segregating Non-Syndromic Hearing Loss.Int J Mol Sci. 2025 Apr 3;26(7):3337. doi: 10.3390/ijms26073337. Int J Mol Sci. 2025. PMID: 40244166 Free PMC article.
-
Tricellulin Expression and its Deletion Effects in the Endolymphatic Sac.J Int Adv Otol. 2018 Aug;14(2):312-316. doi: 10.5152/iao.2018.5473. J Int Adv Otol. 2018. PMID: 30100545 Free PMC article.
-
Transgenic animal models to explore and modulate the blood brain and blood retinal barriers of the CNS.Fluids Barriers CNS. 2022 Nov 1;19(1):86. doi: 10.1186/s12987-022-00386-0. Fluids Barriers CNS. 2022. PMID: 36320068 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
