

Selective Inhibition of the Mitochondrial Permeability Transition Pore Protects against Neurodegeneration in Experimental Multiple Sclerosis
- PMID: 26679998
- PMCID: PMC4813465
- DOI: 10.1074/jbc.M115.700385
Selective Inhibition of the Mitochondrial Permeability Transition Pore Protects against Neurodegeneration in Experimental Multiple Sclerosis
Abstract
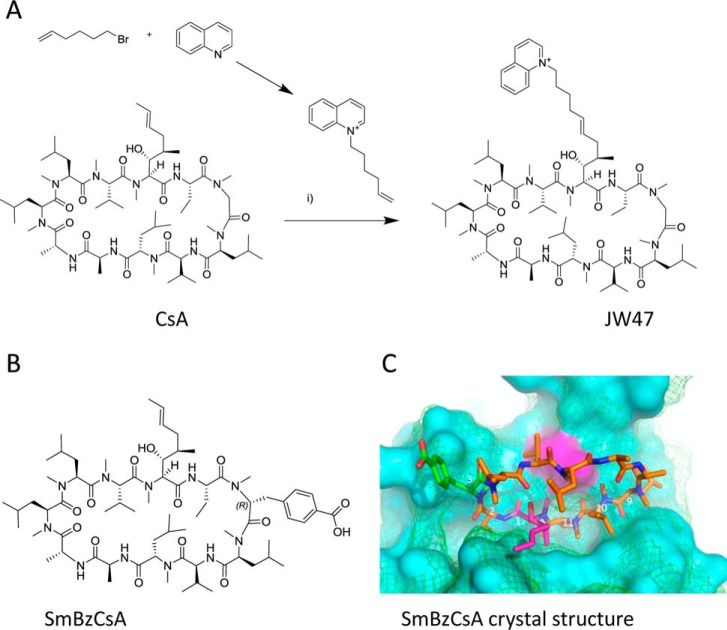
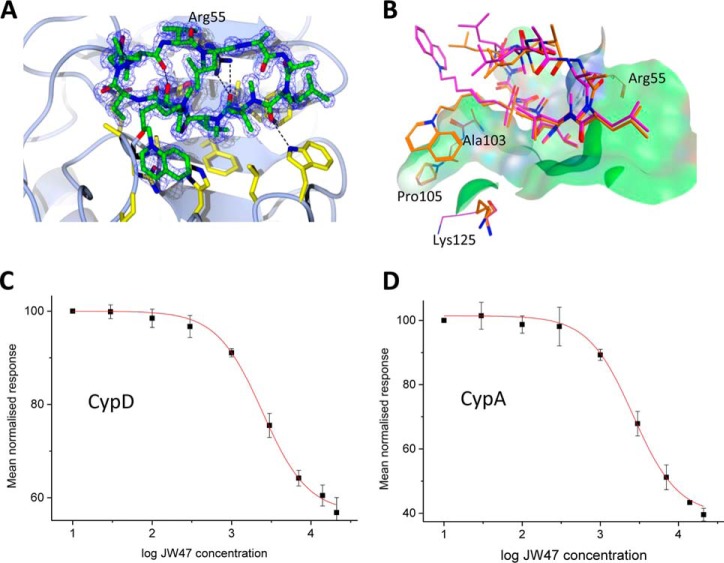
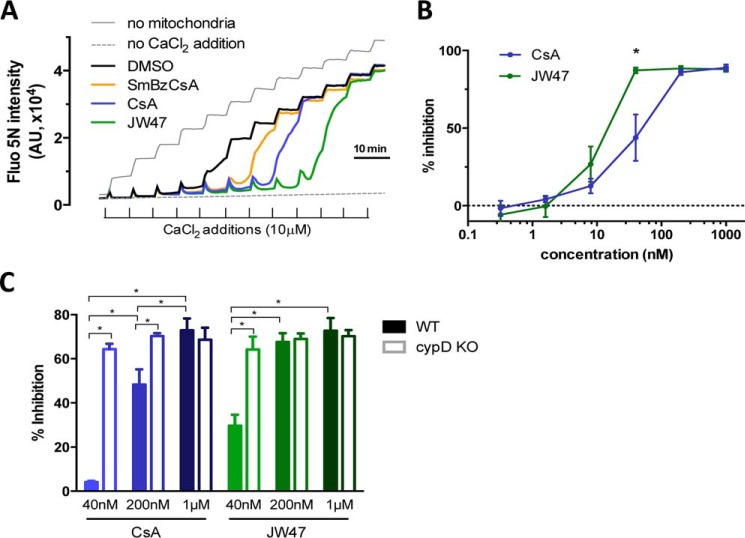
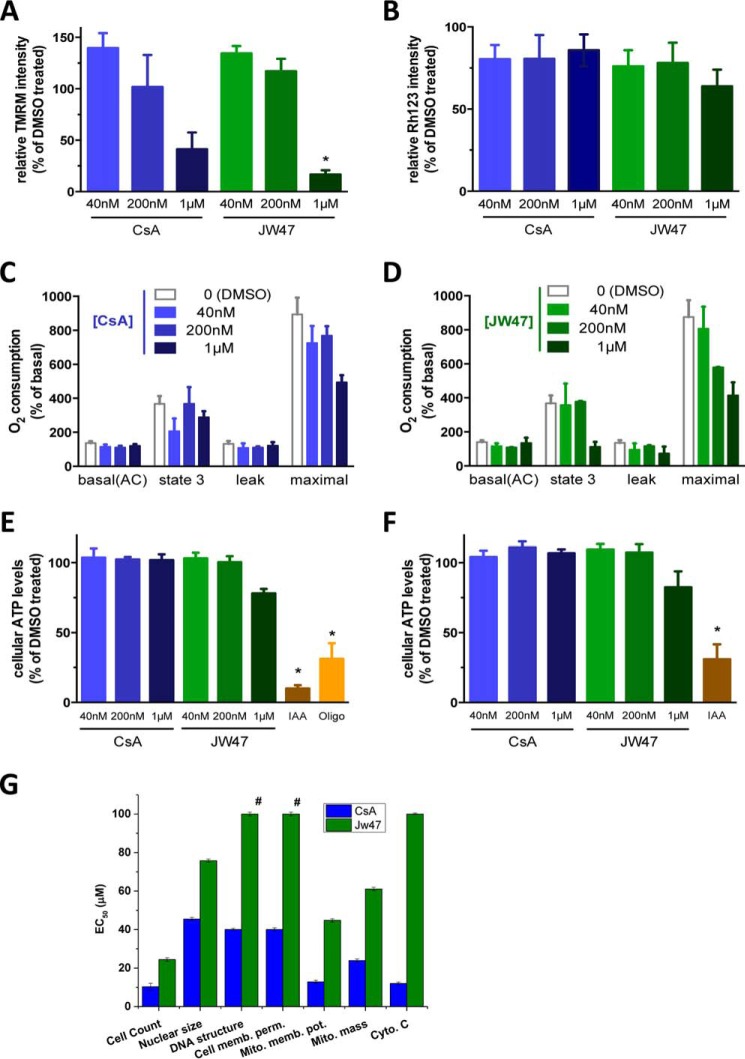
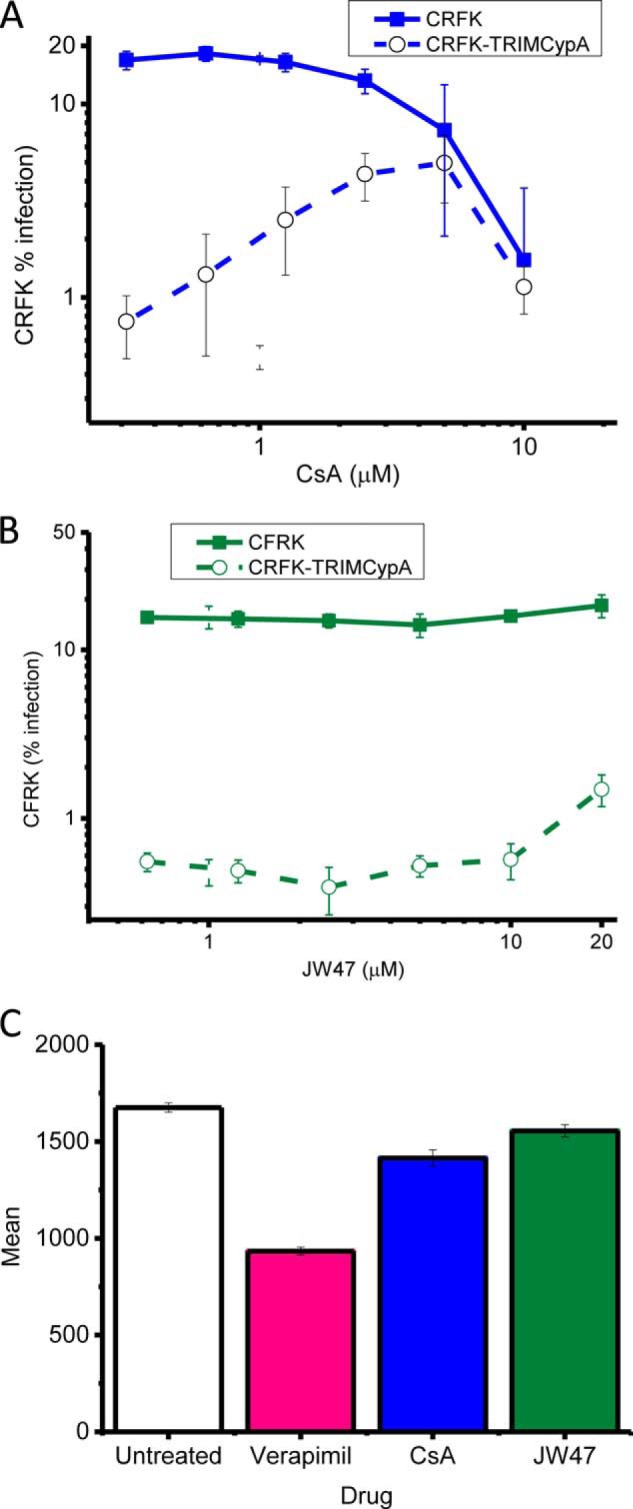
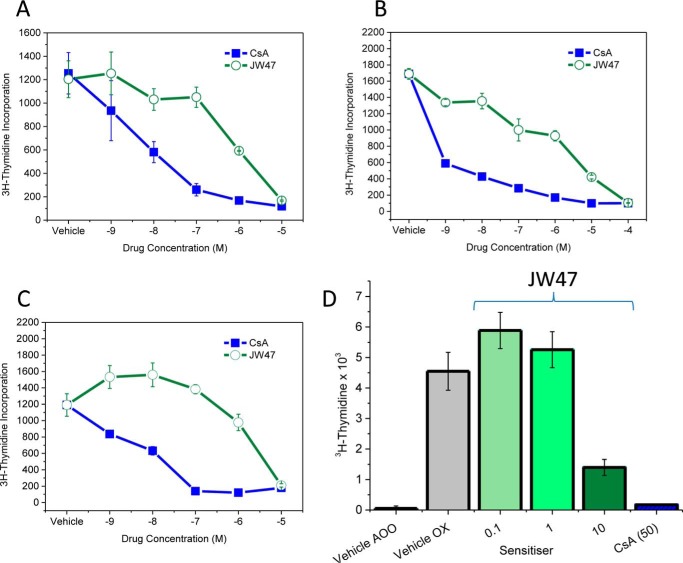
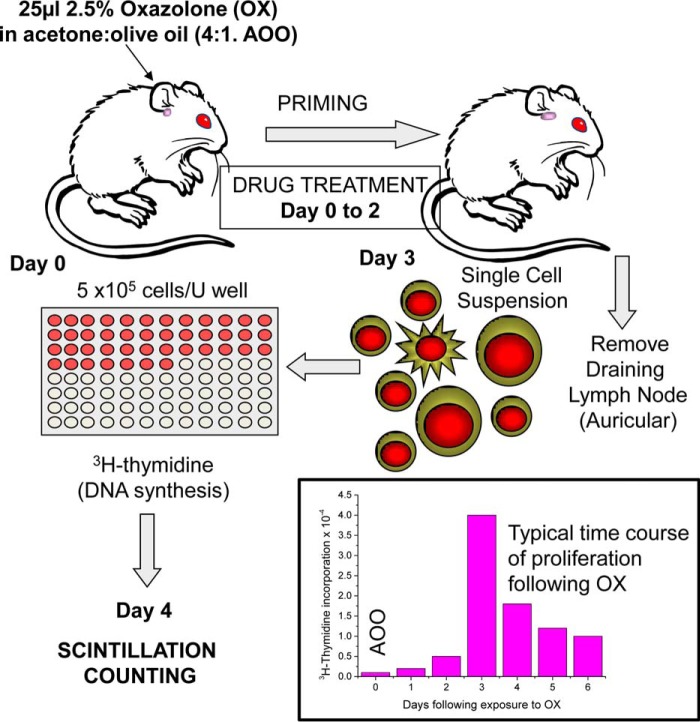
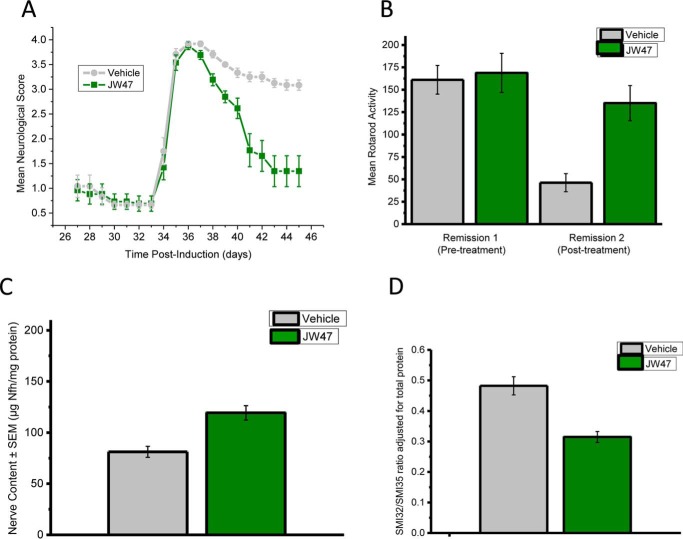
The mitochondrial permeability transition pore is a recognized drug target for neurodegenerative conditions such as multiple sclerosis and for ischemia-reperfusion injury in the brain and heart. The peptidylprolyl isomerase, cyclophilin D (CypD, PPIF), is a positive regulator of the pore, and genetic down-regulation or knock-out improves outcomes in disease models. Current inhibitors of peptidylprolyl isomerases show no selectivity between the tightly conserved cyclophilin paralogs and exhibit significant off-target effects, immunosuppression, and toxicity. We therefore designed and synthesized a new mitochondrially targeted CypD inhibitor, JW47, using a quinolinium cation tethered to cyclosporine. X-ray analysis was used to validate the design concept, and biological evaluation revealed selective cellular inhibition of CypD and the permeability transition pore with reduced cellular toxicity compared with cyclosporine. In an experimental autoimmune encephalomyelitis disease model of neurodegeneration in multiple sclerosis, JW47 demonstrated significant protection of axons and improved motor assessments with minimal immunosuppression. These findings suggest that selective CypD inhibition may represent a viable therapeutic strategy for MS and identify quinolinium as a mitochondrial targeting group for in vivo use.
Keywords: EAE; X-ray crystallography; cyclophilin D; cyclosporin; mitochondrial permeability transition (MPT); mitochondrial targeting; multiple sclerosis; neurodegeneration; neurodegenerative disease.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Alavian K. N., Beutner G., Lazrove E., Sacchetti S., Park H. A., Licznerski P., Li H., Nabili P., Hockensmith K., Graham M., Porter G. A. Jr., and Jonas E. A. (2014) An uncoupling channel within the c-subunit ring of the F1FO ATP synthase is the mitochondrial permeability transition pore. Proc. Natl. Acad. Sci. U.S.A. 111, 10580–10585 - PMC - PubMed
-
- Bonora M., Bononi A., De Marchi E., Giorgi C., Lebiedzinska M., Marchi S., Patergnani S., Rimessi A., Suski J. M., Wojtala A., Wieckowski M. R., Kroemer G., Galluzzi L., and Pinton P. (2013) Role of the c subunit of the FO ATP synthase in mitochondrial permeability transition. Cell Cycle 12, 674–683 - PMC - PubMed
-
- Halestrap A. P., and Richardson A. P. (2015) The mitochondrial permeability transition: a current perspective on its identity and role in ischaemia/reperfusion injury. J. Mol. Cell. Cardiol. 78, 129–141 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
