

A novel ATAC-seq approach reveals lineage-specific reinforcement of the open chromatin landscape via cooperation between BAF and p63
- PMID: 26683334
- PMCID: PMC4699366
- DOI: 10.1186/s13059-015-0840-9
A novel ATAC-seq approach reveals lineage-specific reinforcement of the open chromatin landscape via cooperation between BAF and p63
Abstract
Background: Open chromatin regions are correlated with active regulatory elements in development and are dysregulated in diseases. The BAF (SWI/SNF) complex is essential for development, and has been demonstrated to remodel reconstituted chromatin in vitro and to control the accessibility of a few individual regions in vivo. However, it remains unclear where and how BAF controls the open chromatin landscape to regulate developmental processes, such as human epidermal differentiation.
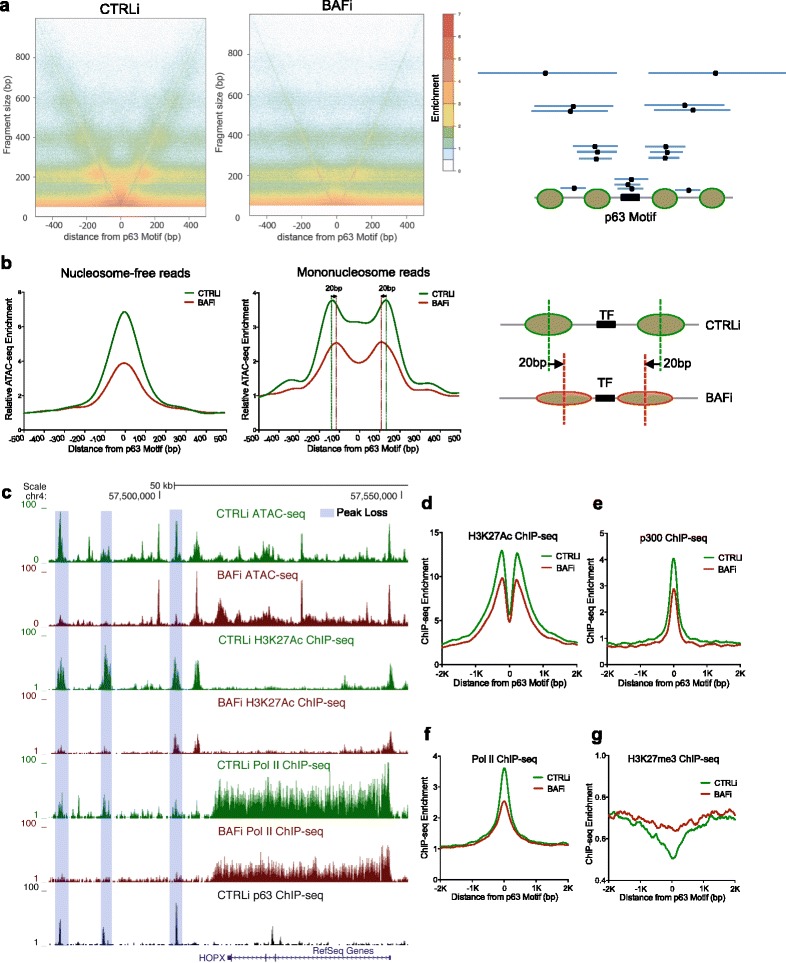
Results: Using a novel "on-plate" ATAC-sequencing approach for profiling open chromatin landscapes with a low number of adherent cells, we demonstrate that the BAF complex is essential for maintaining 11.6 % of open chromatin regions in epidermal differentiation. These BAF-dependent open chromatin regions are highly cell-type-specific and are strongly enriched for binding sites for p63, a master epidermal transcription factor. The DNA sequences of p63 binding sites intrinsically favor nucleosome formation and are inaccessible in other cell types without p63 to prevent ectopic activation. In epidermal cells, BAF and p63 mutually recruit each other to maintain 14,853 open chromatin regions. We further demonstrate that BAF and p63 cooperatively position nucleosomes away from p63 binding sites and recruit transcriptional machinery to control tissue differentiation.
Conclusions: BAF displays high specificity in controlling the open chromatin landscape during epidermal differentiation by cooperating with the master transcription factor p63 to maintain lineage-specific open chromatin regions.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
