

Soybean Aphid Infestation Induces Changes in Fatty Acid Metabolism in Soybean
- PMID: 26684003
- PMCID: PMC4684210
- DOI: 10.1371/journal.pone.0145660
Soybean Aphid Infestation Induces Changes in Fatty Acid Metabolism in Soybean
Abstract
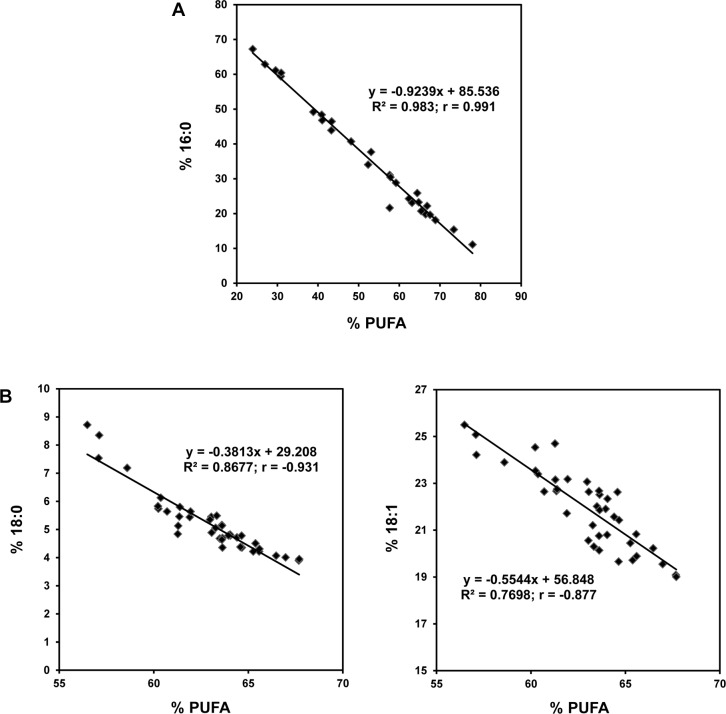
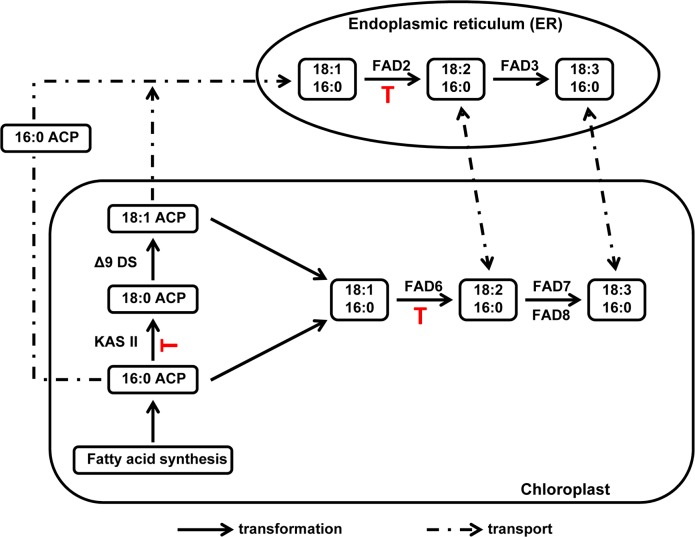
The soybean aphid (Aphis glycines Matsumura) is one of the most important insect pests of soybeans in the North-central region of the US. It has been hypothesized that aphids avoid effective defenses by inhibition of jasmonate-regulated plant responses. Given the role fatty acids play in jasmonate-induced plant defenses, we analyzed the fatty acid profile of soybean leaves and seeds from aphid-infested plants. Aphid infestation reduced levels of polyunsaturated fatty acids in leaves with a concomitant increase in palmitic acid. In seeds, a reduction in polyunsaturated fatty acids was associated with an increase in stearic acid and oleic acid. Soybean plants challenged with the brown stem rot fungus or with soybean cyst nematodes did not present changes in fatty acid levels in leaves or seeds, indicating that the changes induced by aphids are not a general response to pests. One of the polyunsaturated fatty acids, linolenic acid, is the precursor of jasmonate; thus, these changes in fatty acid metabolism may be examples of "metabolic hijacking" by the aphid to avoid the induction of effective defenses. Based on the changes in fatty acid levels observed in seeds and leaves, we hypothesize that aphids potentially induce interference in the fatty acid desaturation pathway, likely reducing FAD2 and FAD6 activity that leads to a reduction in polyunsaturated fatty acids. Our data support the idea that aphids block jasmonate-dependent defenses by reduction of the hormone precursor.
Conflict of interest statement
Figures
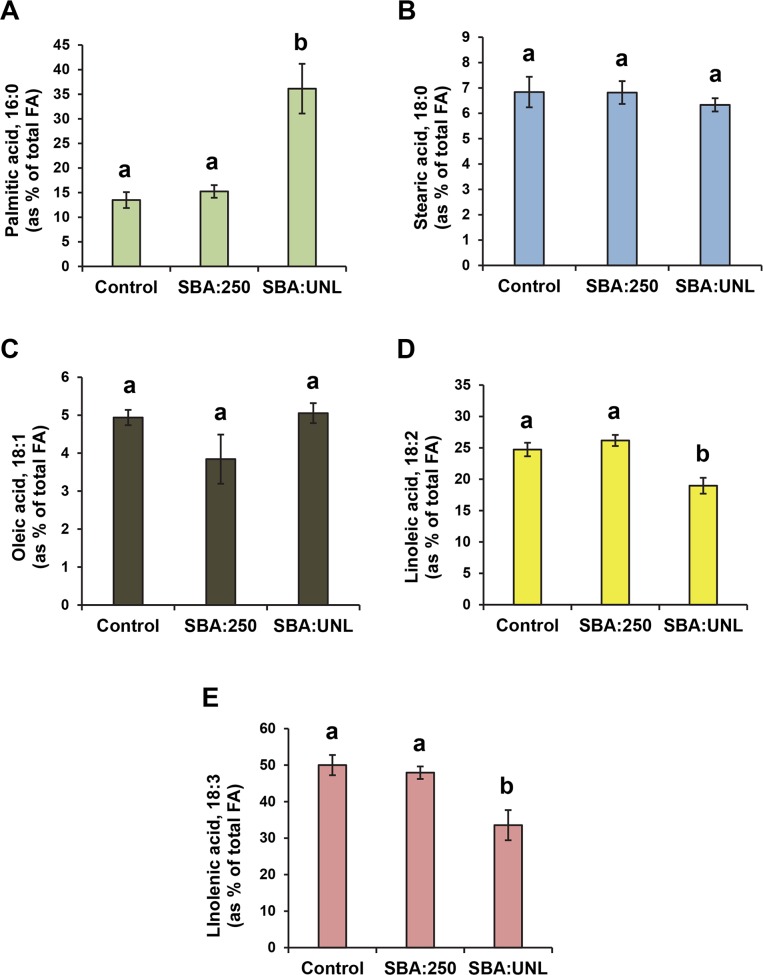
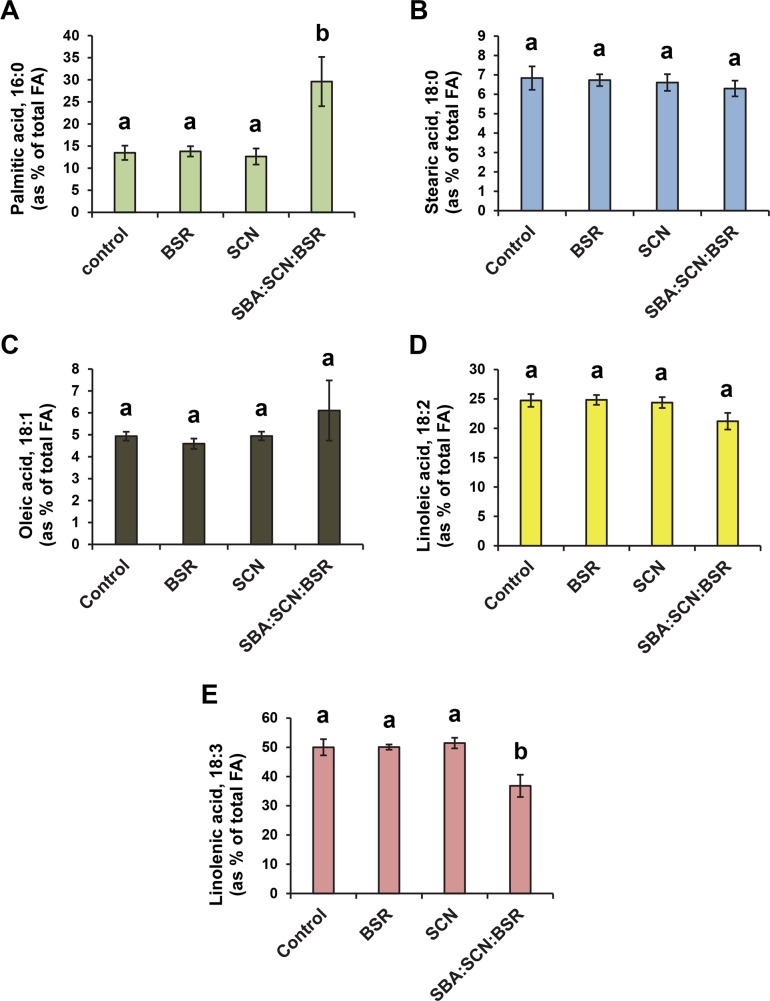
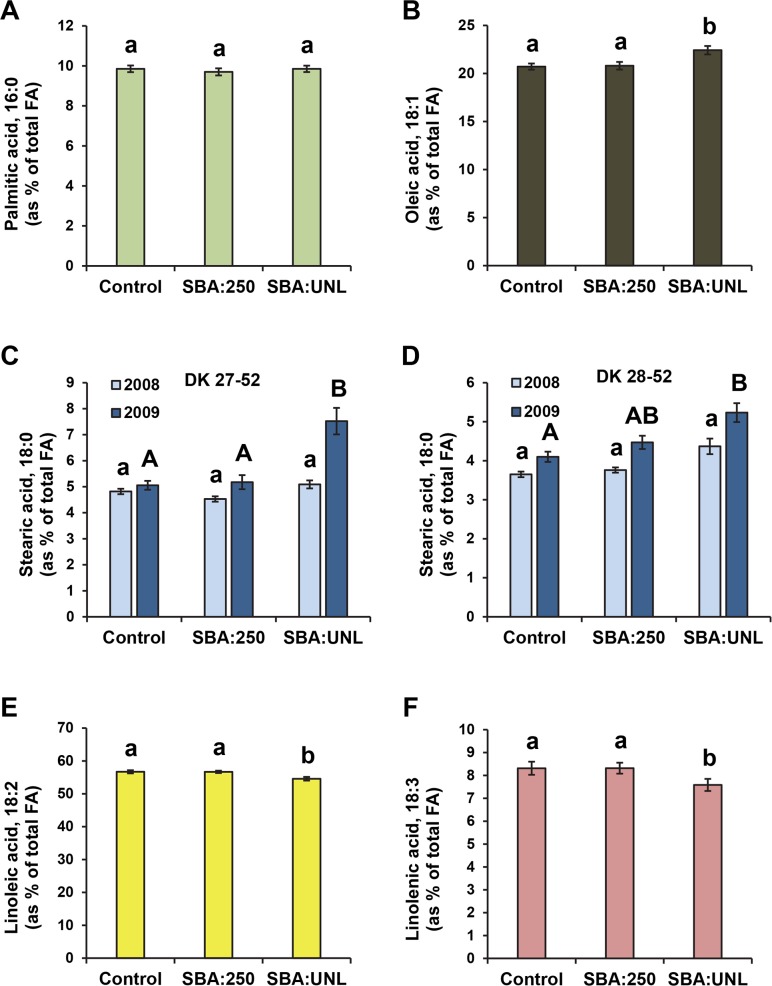
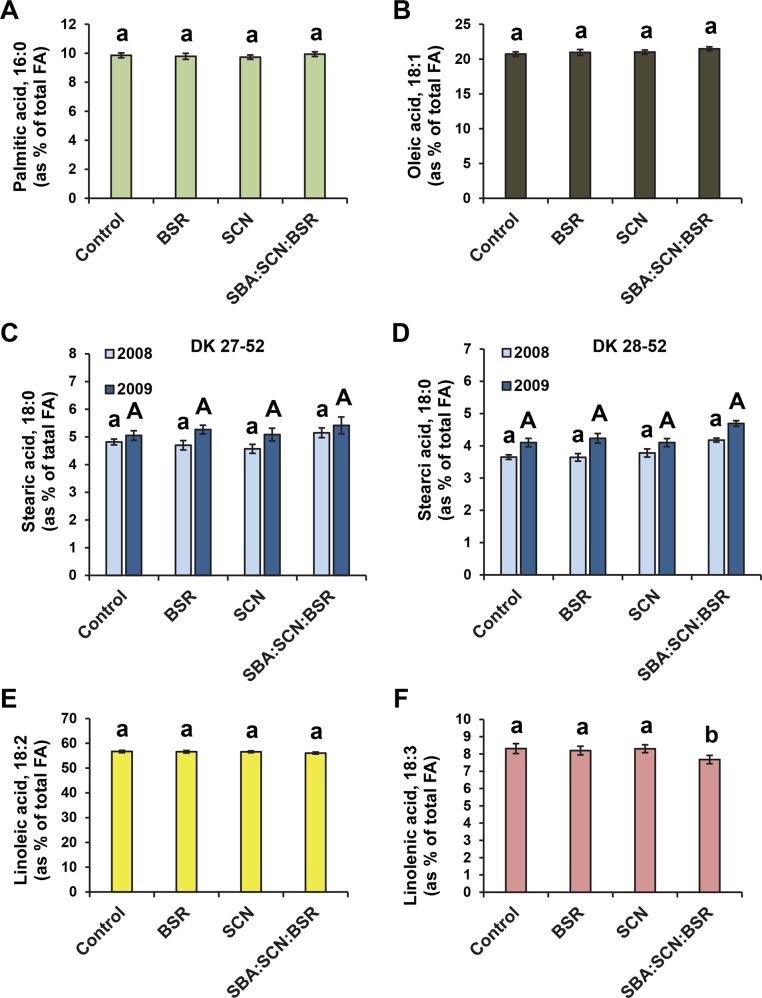
References
-
- Grant M, Lamb C. Systemic immunity. Curr Opin Plant Biol. 2006;9(4):414–420. - PubMed
-
- Howe GA. Jasmonates as signals in the wound response. J Plant Growth Regul. 2004;23(3):223–237.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
