

Identification of the Post-translational Modifications Present in Centromeric Chromatin
- PMID: 26685127
- PMCID: PMC4813710
- DOI: 10.1074/mcp.M115.053710
Identification of the Post-translational Modifications Present in Centromeric Chromatin
Abstract
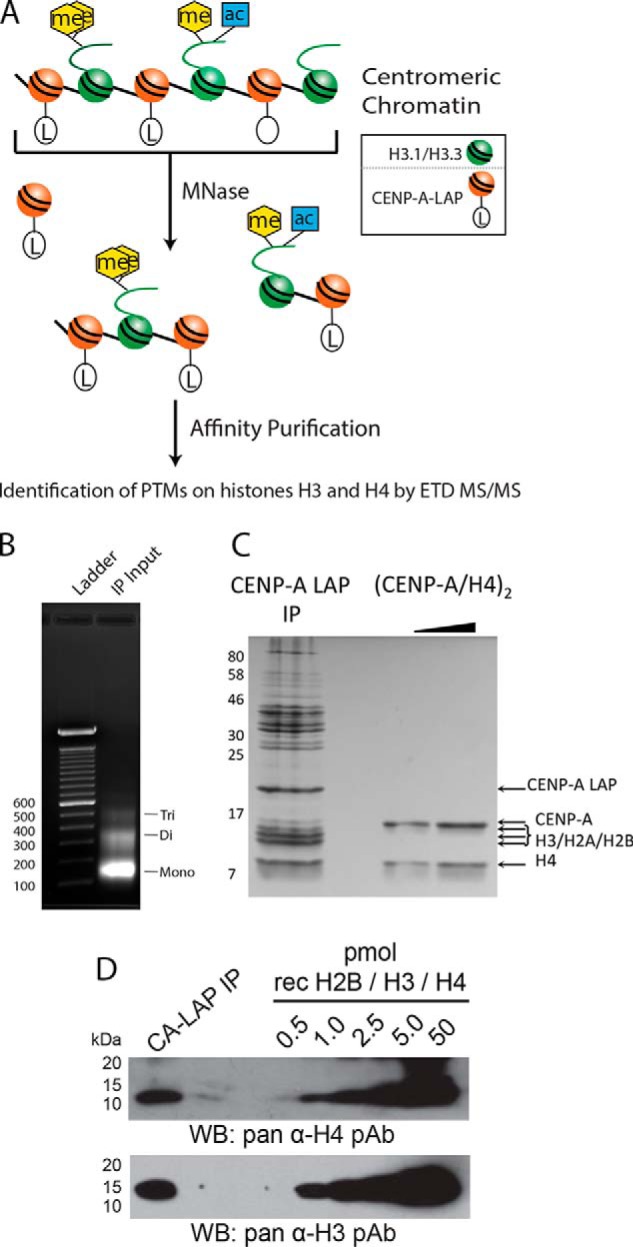
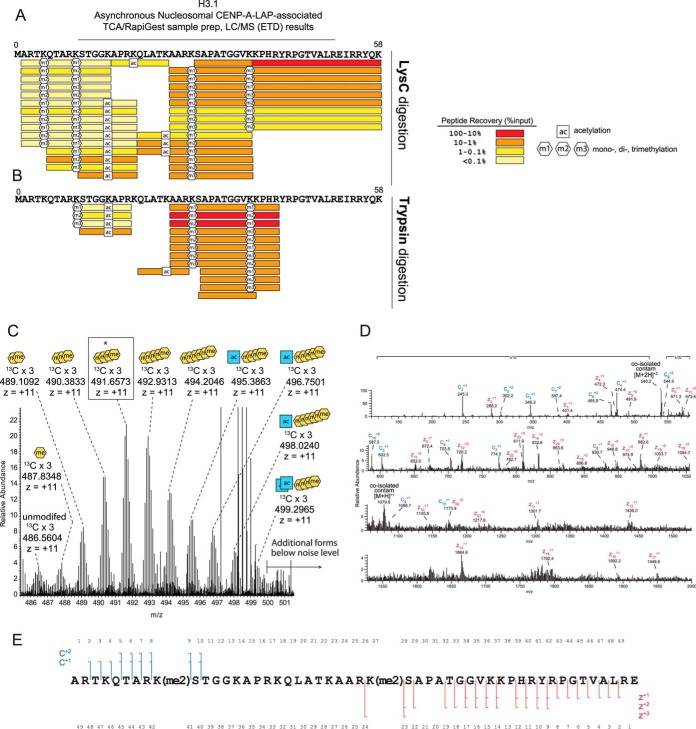
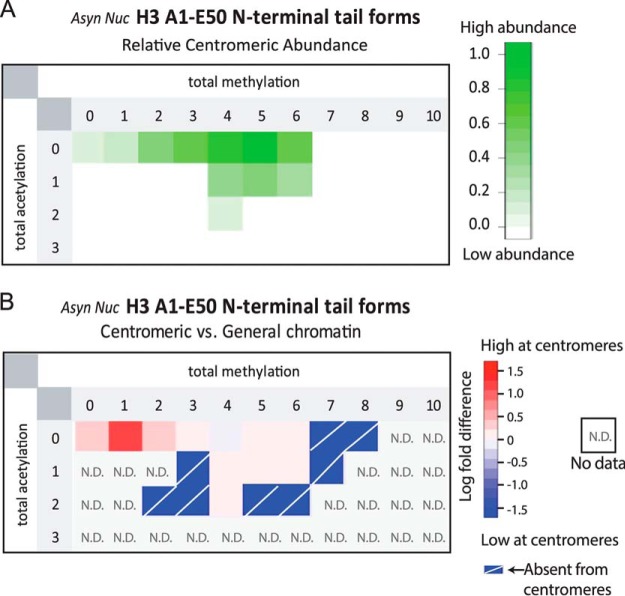
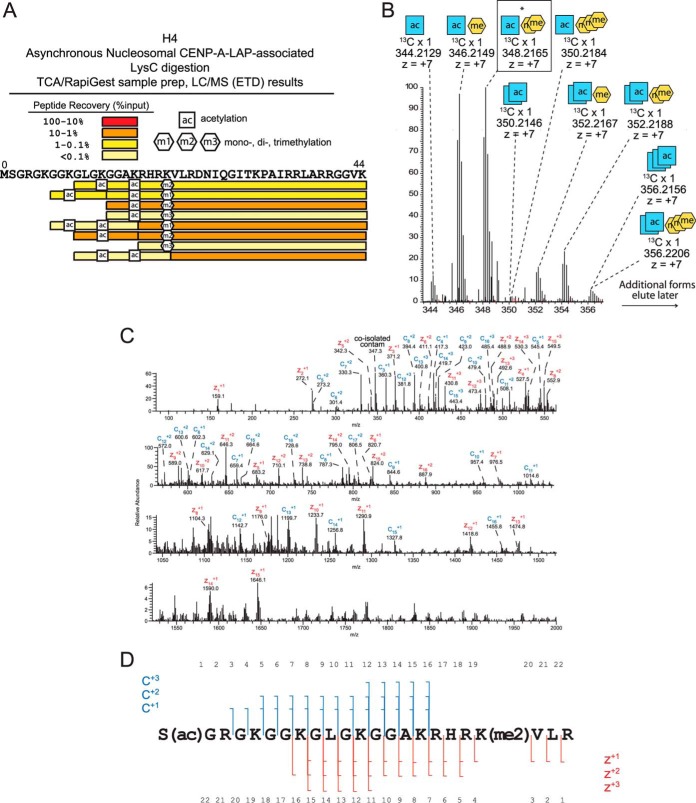
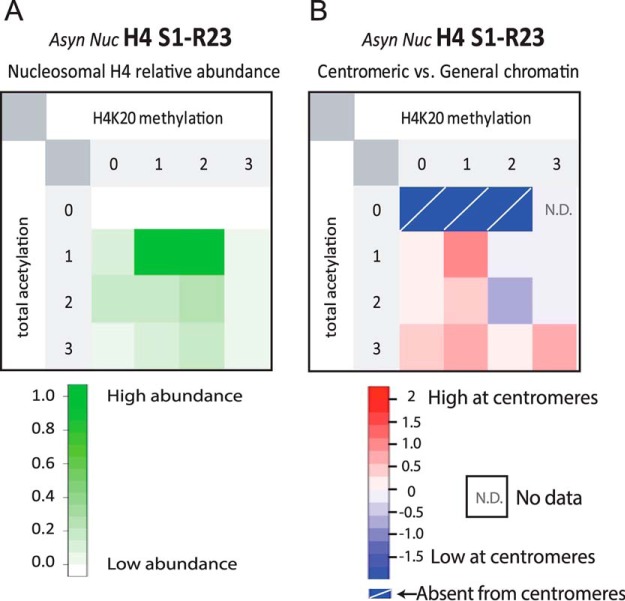
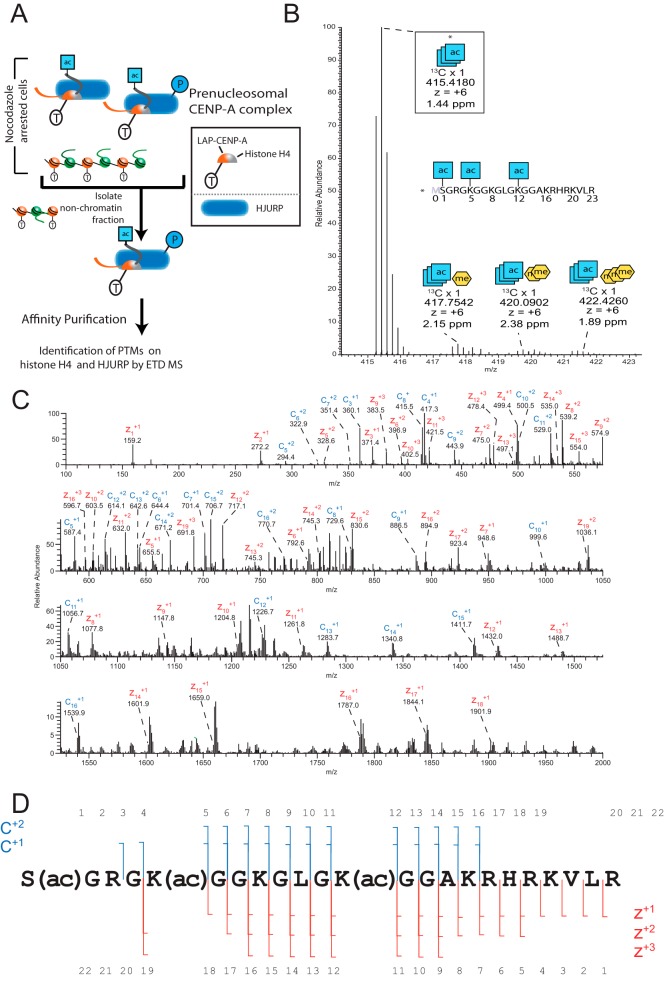
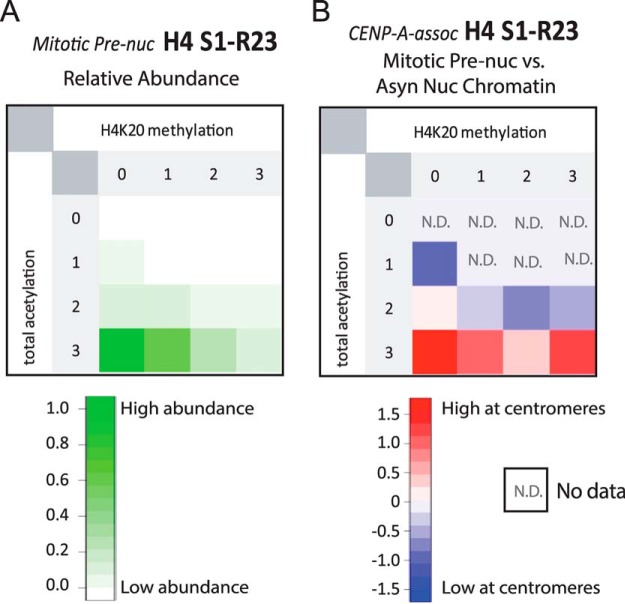
The centromere is the locus on the chromosome that acts as the essential connection point between the chromosome and the mitotic spindle. A histone H3 variant, CENP-A, defines the location of the centromere, but centromeric chromatin consists of a mixture of both CENP-A-containing and H3-containing nucleosomes. We report a surprisingly uniform pattern of primarily monomethylation on lysine 20 of histone H4 present in short polynucleosomes mixtures of CENP-A and H3 nucleosomes isolated from functional centromeres. Canonical H3 is not a component of CENP-A-containing nucleosomes at centromeres, so the H3 we copurify from these preparations comes exclusively from adjacent nucleosomes. We find that CENP-A-proximal H3 nucleosomes are not uniformly modified but contain a complex set of PTMs. Dually modified K9me2-K27me2 H3 nucleosomes are observed at the centromere. Side-chain acetylation of both histone H3 and histone H4 is low at the centromere. Prior to assembly at centromeres, newly expressed CENP-A is sequestered for a large portion of the cell cycle (late S-phase, G2, and most of mitosis) in a complex that contains its partner, H4, and its chaperone, HJURP. In contrast to chromatin associated centromeric histone H4, we show that prenucleosomal CENP-A-associated histone H4 lacks K20 methylation and contains side-chain and α-amino acetylation. We show HJURP displays a complex set of serine phosphorylation that may potentially regulate the deposition of CENP-A. Taken together, our findings provide key information regarding some of the key components of functional centromeric chromatin.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
