

Mapping Sub-Second Structure in Mouse Behavior
- PMID: 26687221
- PMCID: PMC4708087
- DOI: 10.1016/j.neuron.2015.11.031
Mapping Sub-Second Structure in Mouse Behavior
Abstract
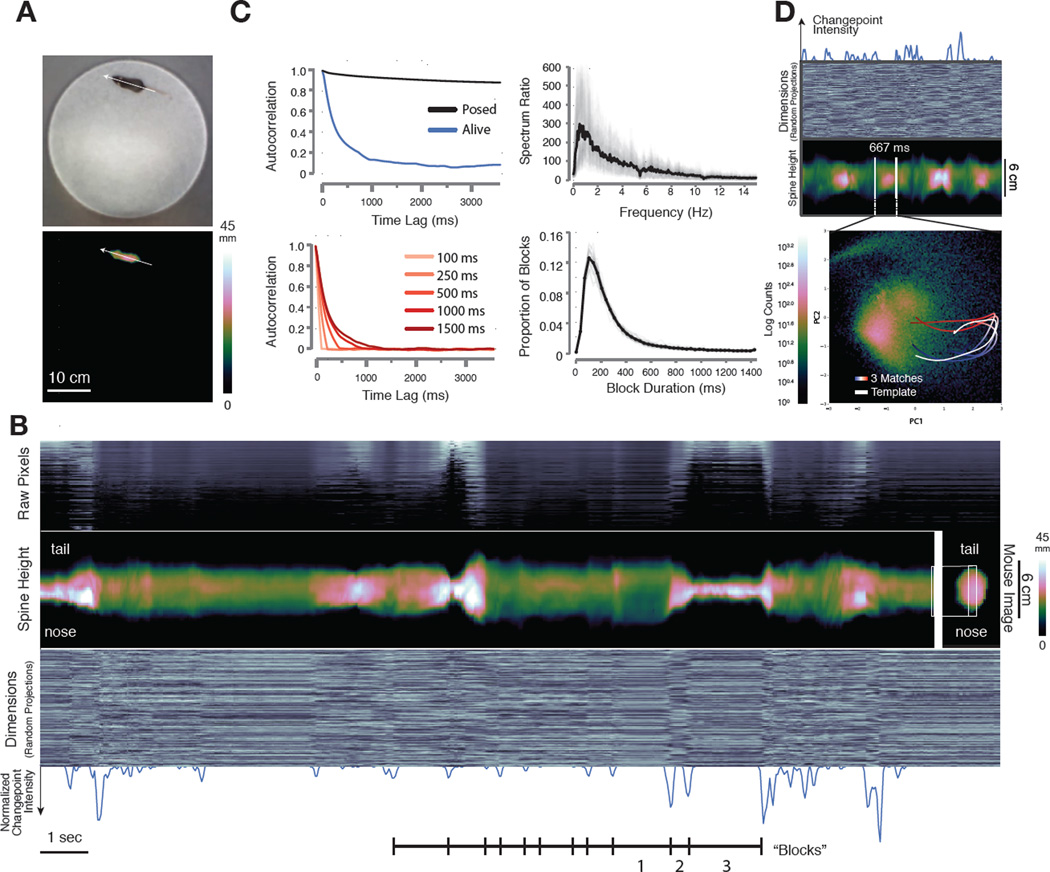
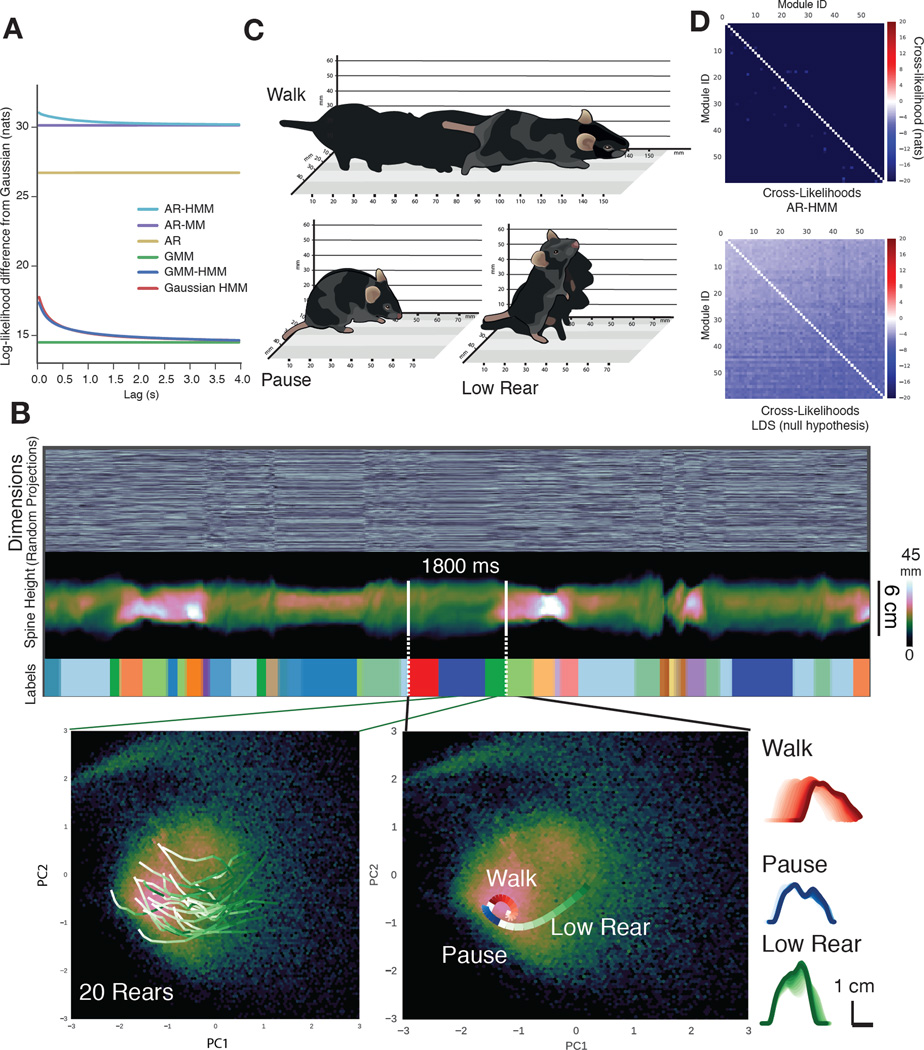
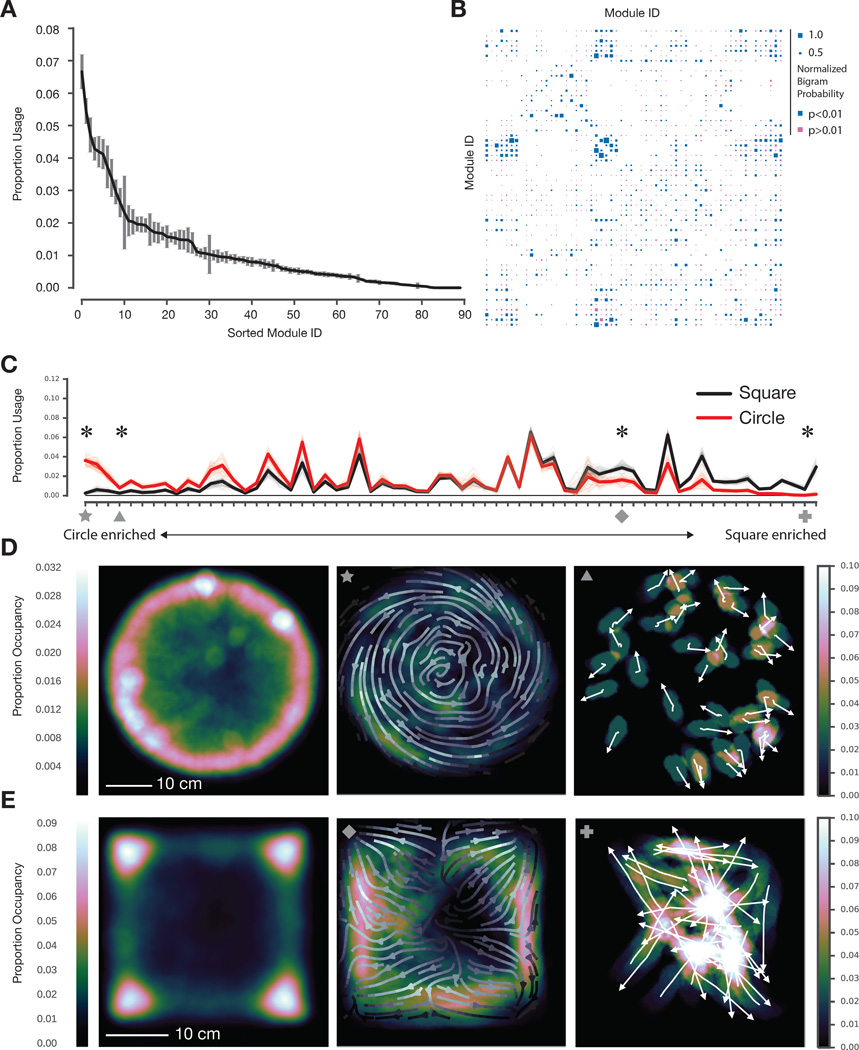
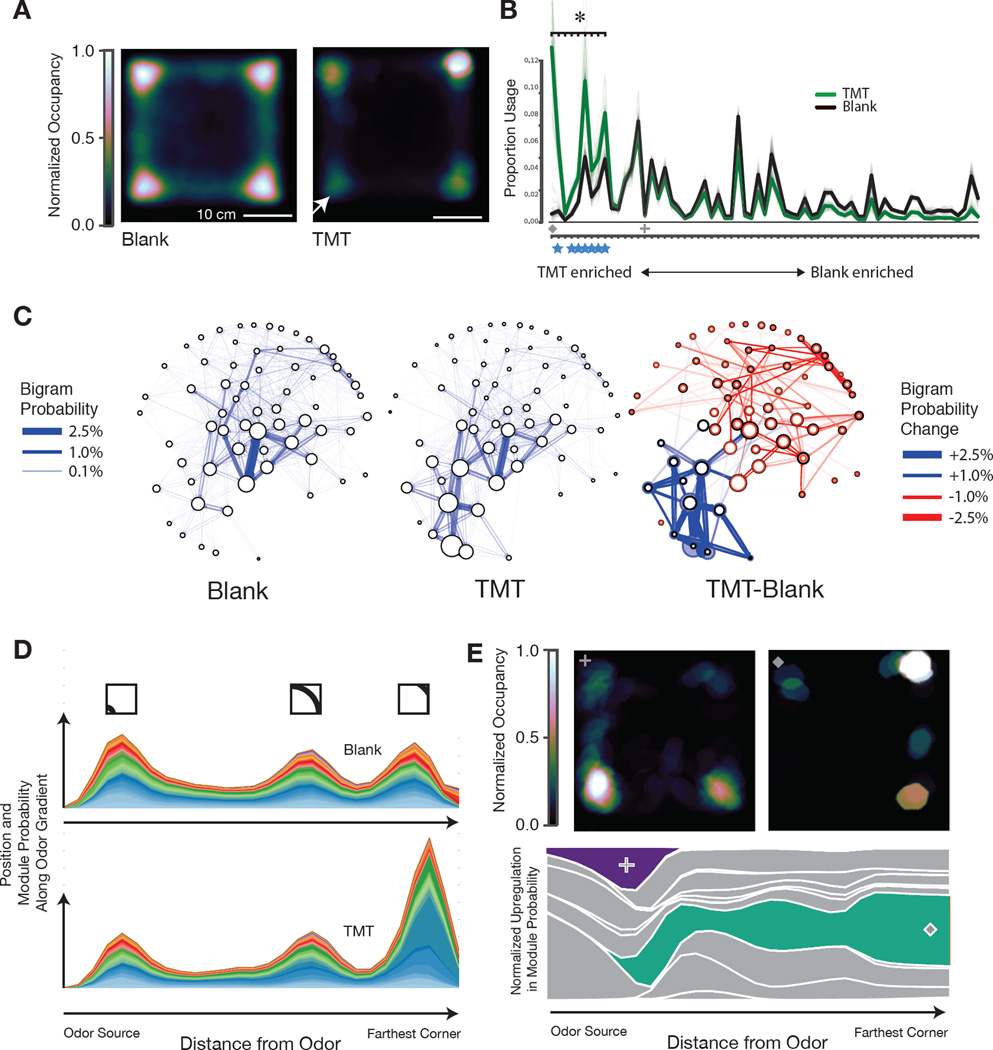
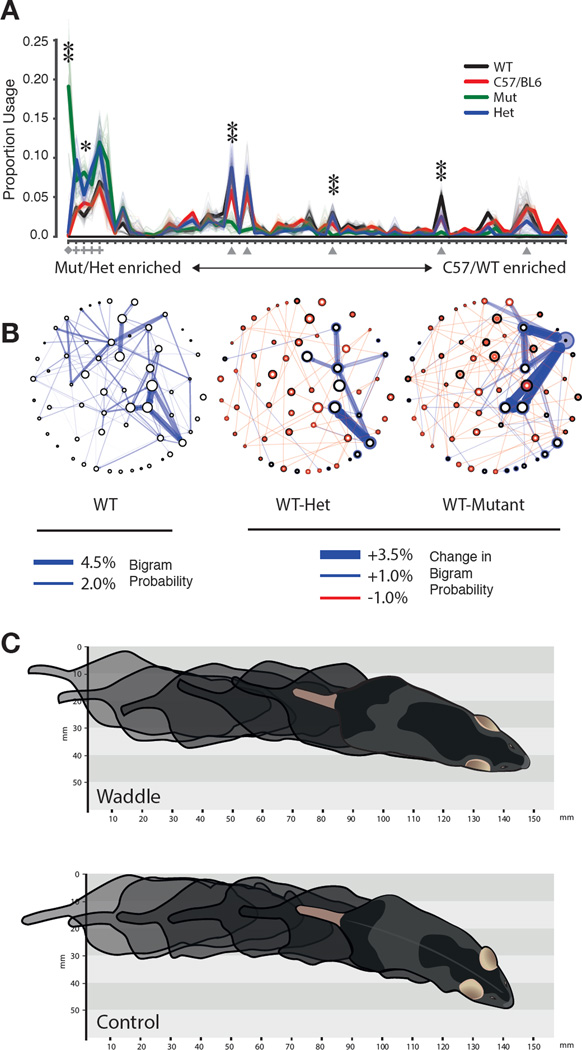
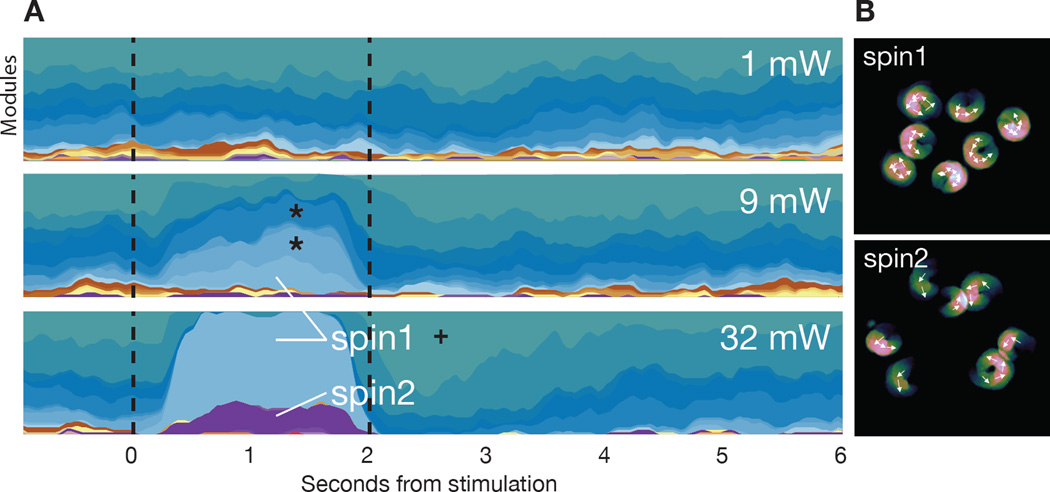
Complex animal behaviors are likely built from simpler modules, but their systematic identification in mammals remains a significant challenge. Here we use depth imaging to show that 3D mouse pose dynamics are structured at the sub-second timescale. Computational modeling of these fast dynamics effectively describes mouse behavior as a series of reused and stereotyped modules with defined transition probabilities. We demonstrate this combined 3D imaging and machine learning method can be used to unmask potential strategies employed by the brain to adapt to the environment, to capture both predicted and previously hidden phenotypes caused by genetic or neural manipulations, and to systematically expose the global structure of behavior within an experiment. This work reveals that mouse body language is built from identifiable components and is organized in a predictable fashion; deciphering this language establishes an objective framework for characterizing the influence of environmental cues, genes and neural activity on behavior.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Aldridge JW, Berridge KC, Rosen AR. Basal ganglia neural mechanisms of natural movement sequences. Canadian Journal of Physiology and Pharmacology. 2004;82:732–739. - PubMed
-
- Anderson DJ, Perona P. Toward a science of computational ethology. Neuron. 2014;84:18–31. - PubMed
-
- Berg HC, Brown DA. Chemotaxis in Escherichia coli analysed by three-dimensional tracking. Nature. 1972;239:500–504. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
