

Glycolate Oxidase Is a Safe and Efficient Target for Substrate Reduction Therapy in a Mouse Model of Primary Hyperoxaluria Type I
- PMID: 26689264
- PMCID: PMC4886931
- DOI: 10.1038/mt.2015.224
Glycolate Oxidase Is a Safe and Efficient Target for Substrate Reduction Therapy in a Mouse Model of Primary Hyperoxaluria Type I
Abstract
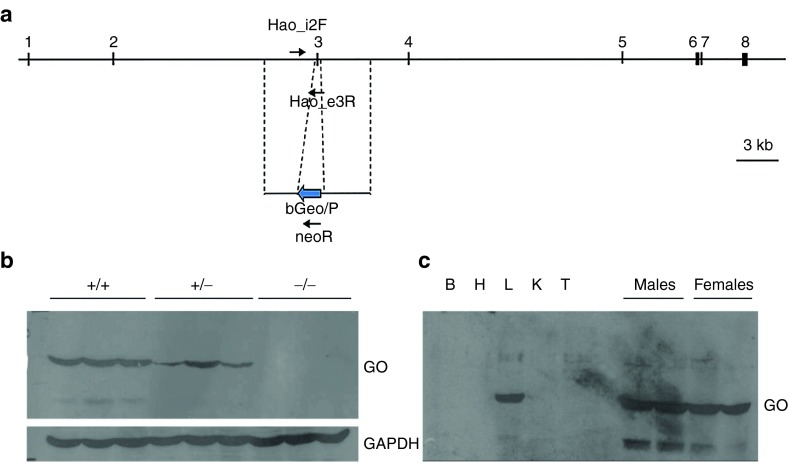
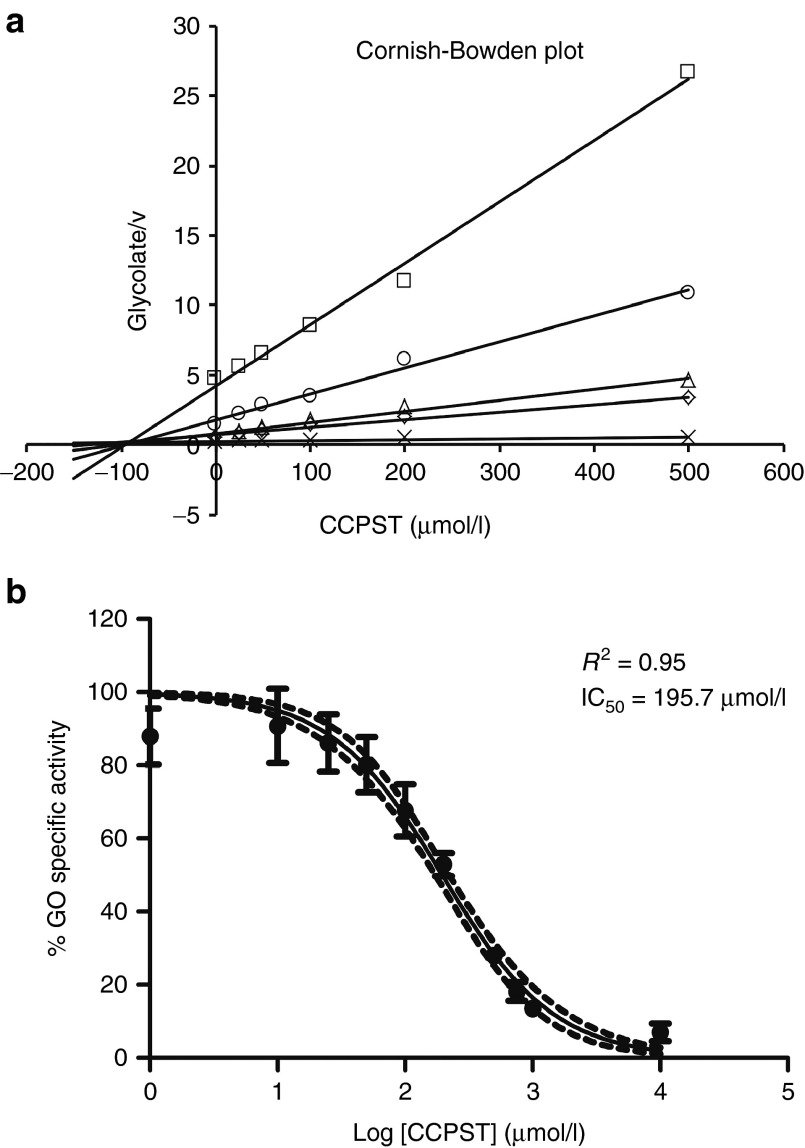
Primary hyperoxaluria type 1 (PH1) is caused by deficient alanine-glyoxylate aminotransferase, the human peroxisomal enzyme that detoxifies glyoxylate. Glycolate is one of the best-known substrates leading to glyoxylate production, via peroxisomal glycolate oxidase (GO). Using genetically modified mice, we herein report GO as a safe and efficient target for substrate reduction therapy (SRT) in PH1. We first generated a GO-deficient mouse (Hao1(-/-)) that presented high urine glycolate levels but no additional phenotype. Next, we produced double KO mice (Agxt1(-/-) Hao1(-/-)) that showed low levels of oxalate excretion compared with hyperoxaluric mice model (Agxt1(-/-)). Previous studies have identified some GO inhibitors, such as 4-carboxy-5-[(4-chlorophenyl)sulfanyl]-1,2,3-thiadiazole (CCPST). We herein report that CCPST inhibits GO in Agxt1(-/-) hepatocytes and significantly reduces their oxalate production, starting at 25 µM. We also tested the ability of orally administered CCPST to reduce oxalate excretion in Agxt1(-/-) mice, showing that 30-50% reduction in urine oxalate can be achieved. In summary, we present proof-of-concept evidence for SRT in PH1. These encouraging results should be followed by a medicinal chemistry programme that might yield more potent GO inhibitors and eventually could result in a pharmacological treatment for this rare and severe inborn error of metabolism.
Figures



Comment in
-
siRNA Therapeutics for Primary Hyperoxaluria: A Beginning.Mol Ther. 2016 Apr;24(4):666-7. doi: 10.1038/mt.2016.50. Mol Ther. 2016. PMID: 27081720 Free PMC article. No abstract available.
-
Re: Glycolate Oxidase is a Safe and Efficient Target for Substrate Reduction Therapy in a Mouse Model of Primary Hyperoxaluria Type I.J Urol. 2016 Jul;196(1):281-2. doi: 10.1016/j.juro.2016.04.005. Epub 2016 Apr 14. J Urol. 2016. PMID: 27321540 No abstract available.
References
-
- Rumsby, G and Cochat, P (2013). Primary hyperoxaluria. N Engl J Med 369: 2163. - PubMed
-
- Salido, E, Pey, AL, Rodriguez, R and Lorenzo, V (2012). Primary hyperoxalurias: disorders of glyoxylate detoxification. Biochim Biophys Acta 1822: 1453–1464. - PubMed
-
- Smid, BE, Aerts, JM, Boot, RG, Linthorst, GE and Hollak, CE (2010). Pharmacological small molecules for the treatment of lysosomal storage disorders. Expert Opin Investig Drugs 19: 1367–1379. - PubMed
-
- Holmes, RP and Assimos, DG (1998). Glyoxylate synthesis, and its modulation and influence on oxalate synthesis. J Urol 160: 1617–1624. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
