

BRET-monitoring of the dynamic changes of inositol lipid pools in living cells reveals a PKC-dependent PtdIns4P increase upon EGF and M3 receptor activation
- PMID: 26692031
- PMCID: PMC4733596
- DOI: 10.1016/j.bbalip.2015.12.005
BRET-monitoring of the dynamic changes of inositol lipid pools in living cells reveals a PKC-dependent PtdIns4P increase upon EGF and M3 receptor activation
Abstract
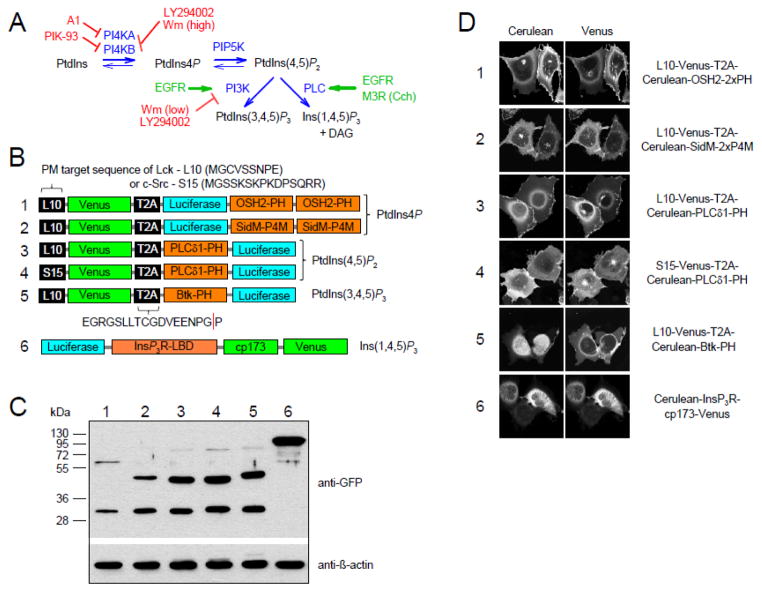
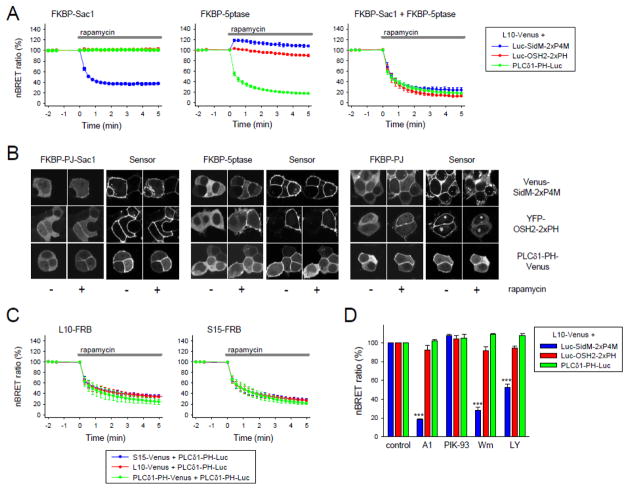
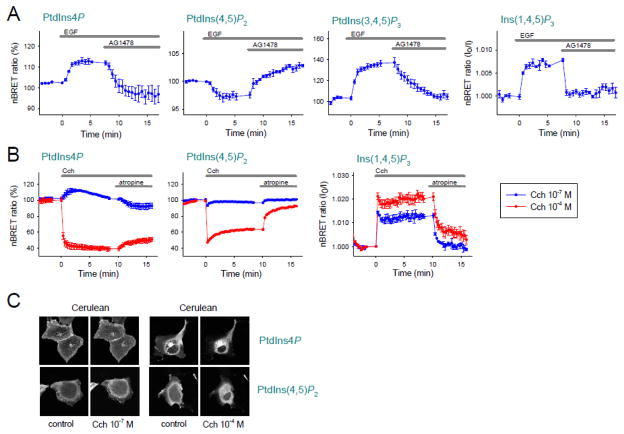
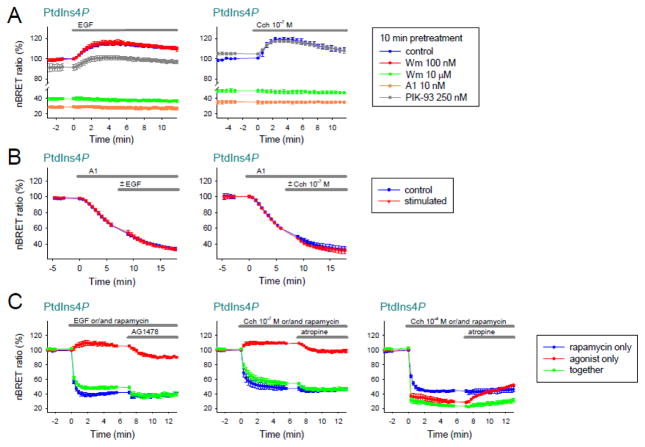
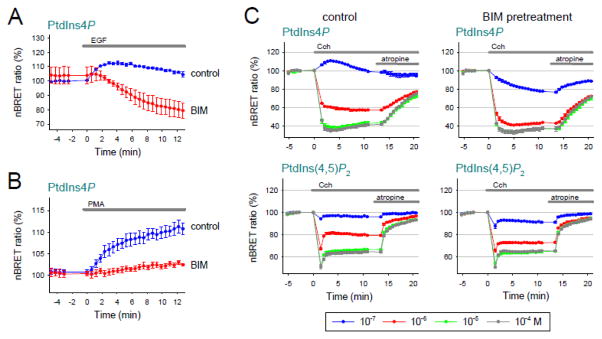
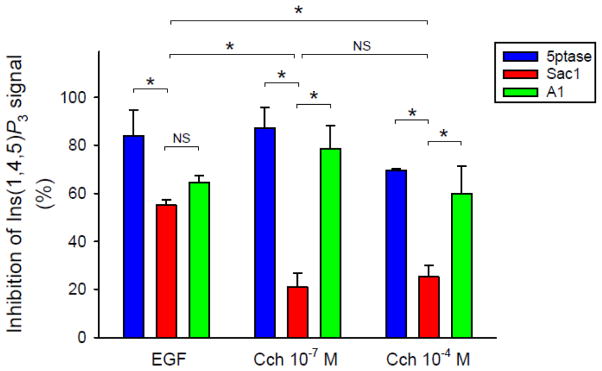
Deciphering many roles played by inositol lipids in signal transduction and membrane function demands experimental approaches that can detect their dynamic accumulation with subcellular accuracy and exquisite sensitivity. The former criterion is met by imaging of fluorescence biosensors in living cells, whereas the latter is facilitated by biochemical measurements from populations. Here, we introduce BRET-based biosensors able to detect rapid changes in inositol lipids in cell populations with both high sensitivity and subcellular resolution in a single, convenient assay. We demonstrate robust and sensitive measurements of PtdIns4P, PtdIns(4,5)P2 and PtdIns(3,4,5)P3 dynamics, as well as changes in cytoplasmic Ins(1,4,5)P3 levels. Measurements were made during either experimental activation of lipid degradation, or PI 3-kinase and phospholipase C mediated signal transduction. Our results reveal a previously unappreciated synthesis of PtdIns4P that accompanies moderate activation of phospholipase C signaling downstream of both EGF and muscarinic M3 receptor activation. This signaling-induced PtdIns4P synthesis relies on protein kinase C, and implicates a feedback mechanism in the control of inositol lipid metabolism during signal transduction.
Keywords: BRET; EGF receptor; GPCR; PI4-kinase; PKC; Phosphoinositides.
Copyright © 2015 Elsevier B.V. All rights reserved.
Figures
References
-
- Sasaki T, Takasuga S, Sasaki J, Kofuji S, Eguchi S, Yamazaki M, Suzuki A. Mammalian phosphoinositide kinases and phosphatases. Prog Lipid Res. 2009;48:307–343. - PubMed
-
- Di Paolo G, De Camilli P. Phosphoinositides in cell regulation and membrane dynamics. Nature. 2006;443:651–657. - PubMed
-
- Traynor-Kaplan AE, Harris AL, Thompson BL, Taylor P, Sklar LA. An inositol tetrakisphosphate-containing phospholipid in activated neutrophils. Nature. 1988;334:353–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
