

Lysosome associated membrane proteins maintain pancreatic acinar cell homeostasis: LAMP-2 deficient mice develop pancreatitis
- PMID: 26693174
- PMCID: PMC4673685
- DOI: 10.1016/j.jcmgh.2015.07.006
Lysosome associated membrane proteins maintain pancreatic acinar cell homeostasis: LAMP-2 deficient mice develop pancreatitis
Abstract
Background & aims: The pathogenic mechanism of pancreatitis is poorly understood. Recent evidence implicates defective autophagy in pancreatitis responses; however, the pathways mediating impaired autophagy in pancreas remain largely unknown. Here, we investigate the role of lysosome associated membrane proteins (LAMPs) in pancreatitis.
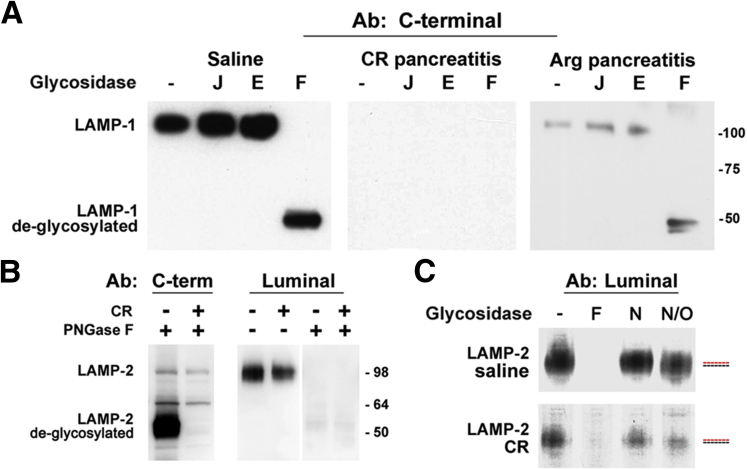
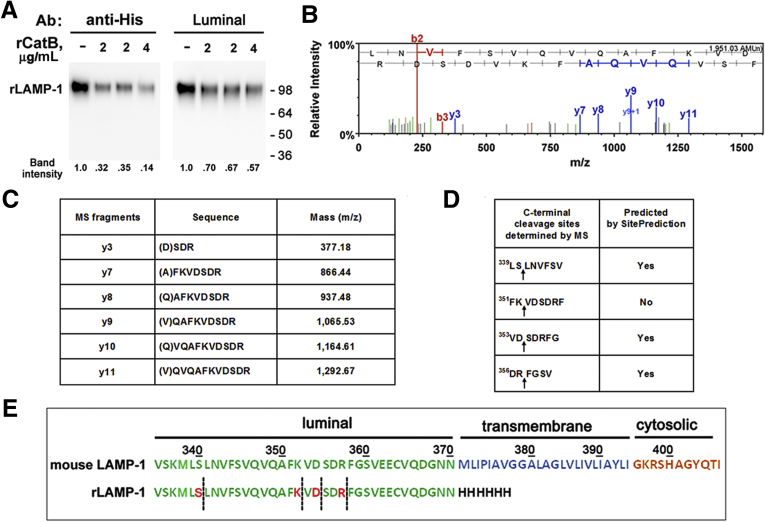
Methods: We analyzed changes in LAMPs in experimental models and human pancreatitis, and the underlying mechanisms: LAMP de-glycosylation and degradation. LAMP cleavage by cathepsin B (CatB) was analyzed by mass spectrometry. We used mice deficient in LAMP-2 to assess its role in pancreatitis.
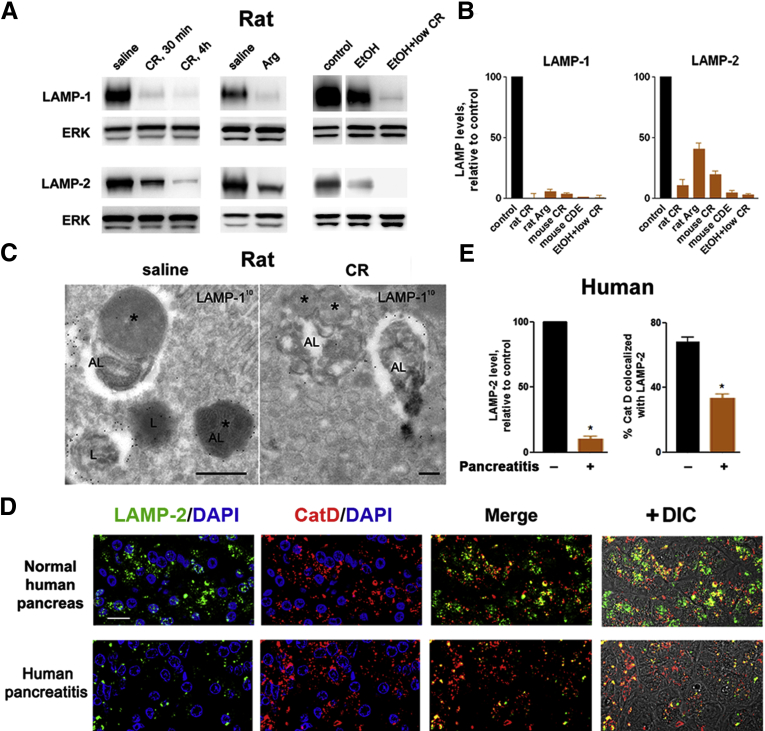
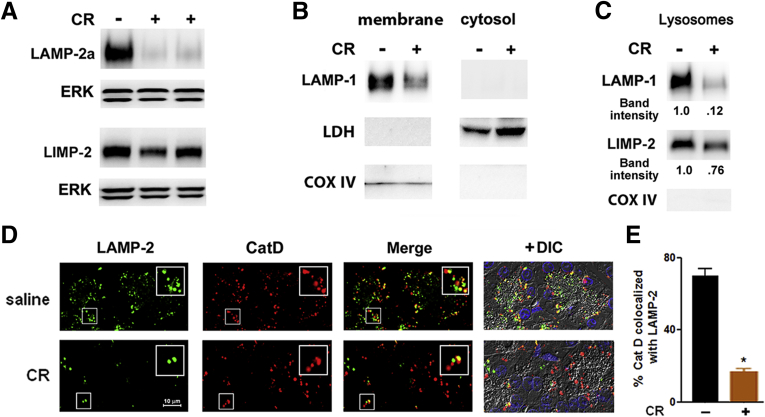
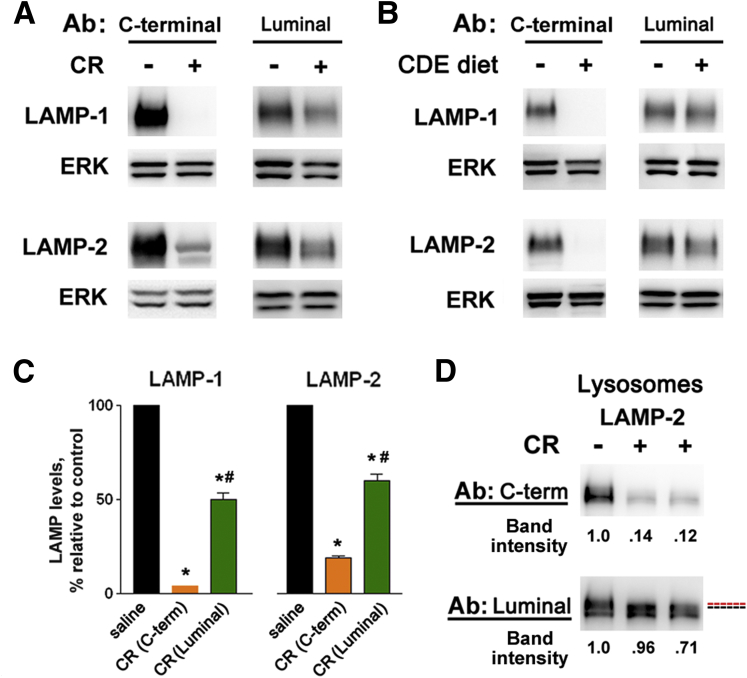
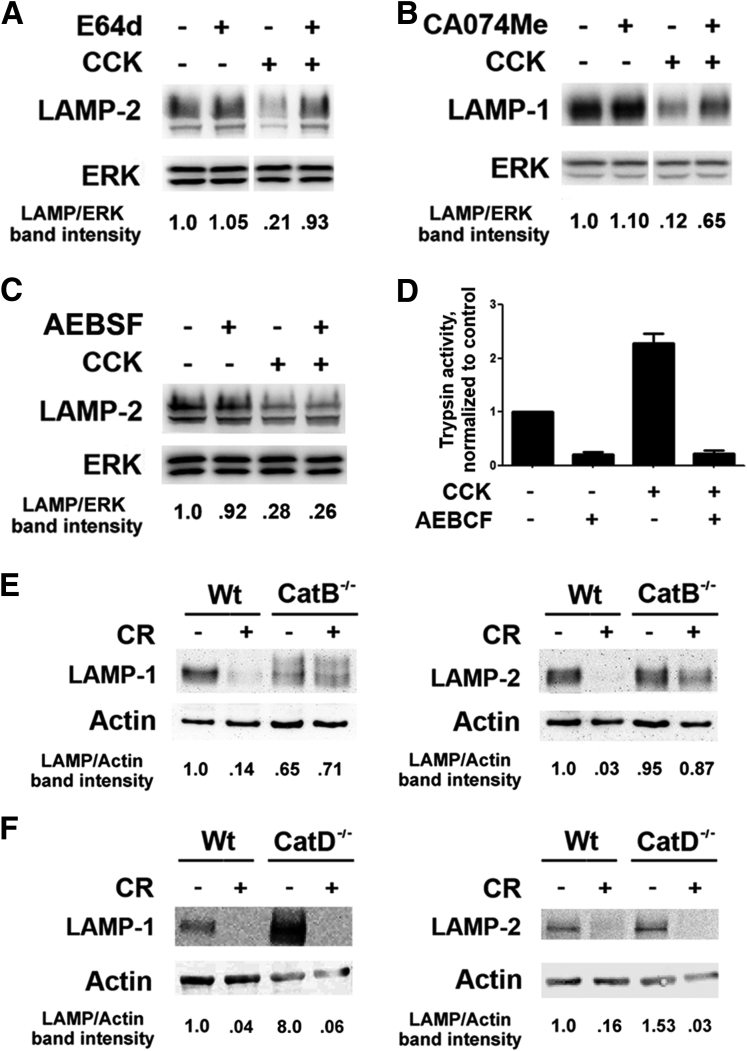
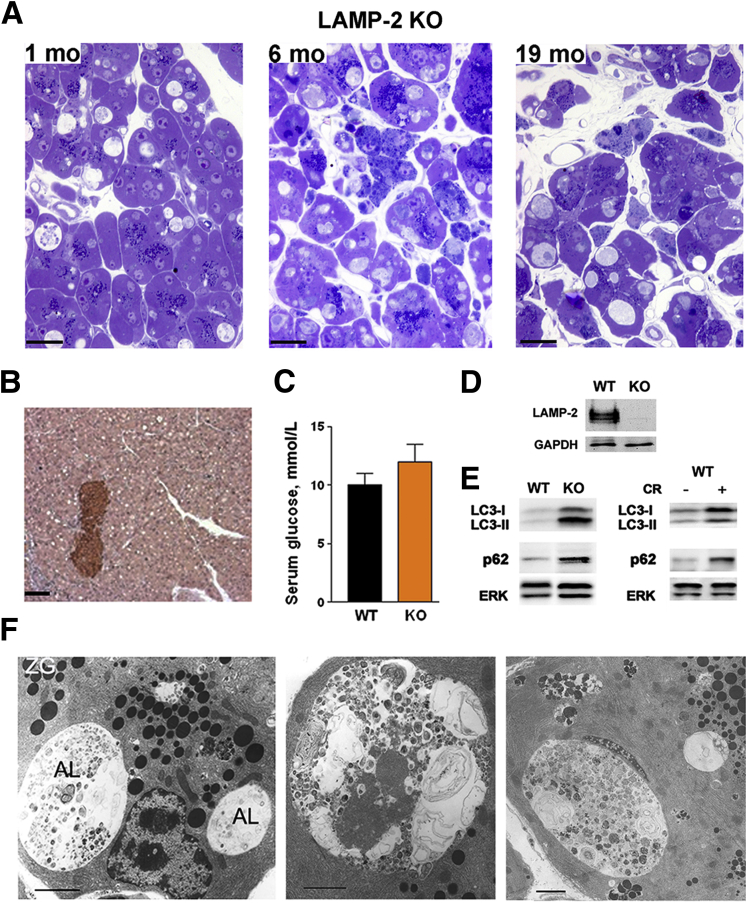
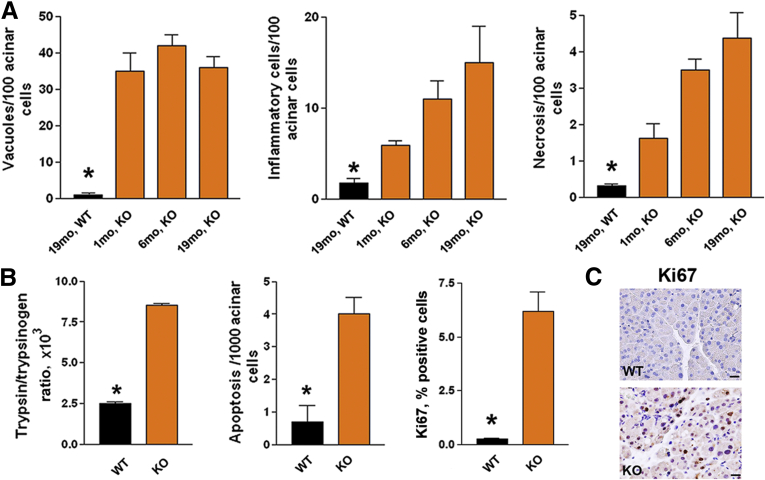
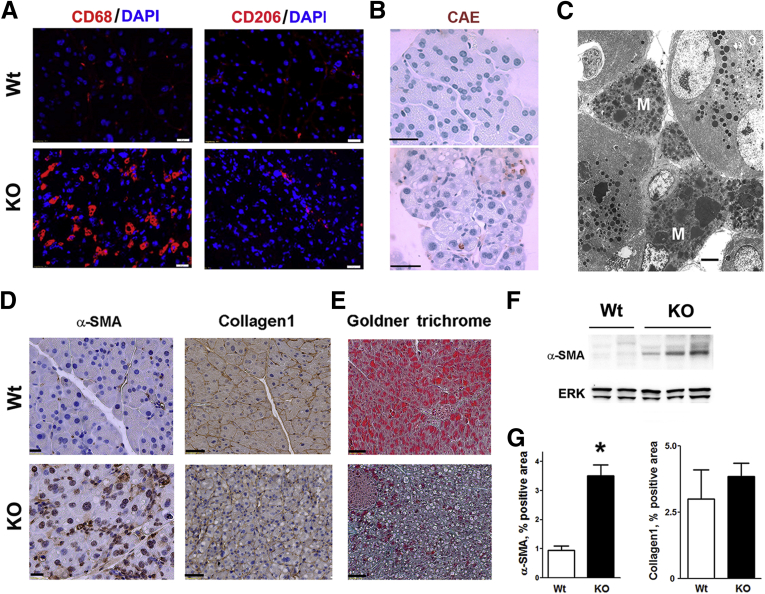
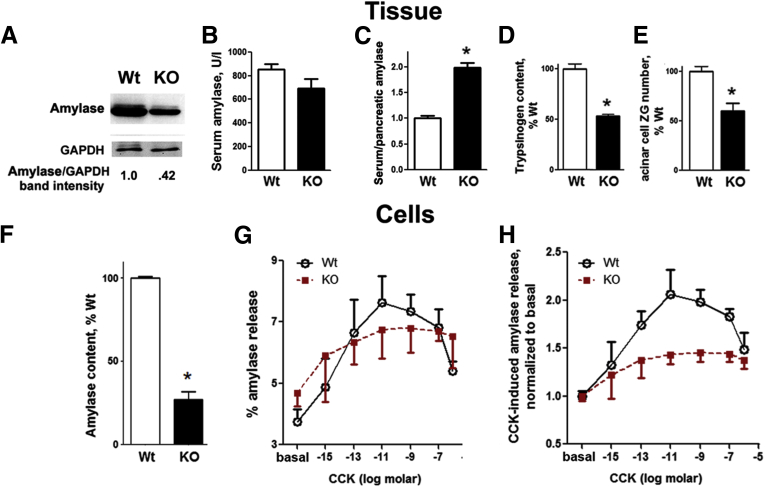
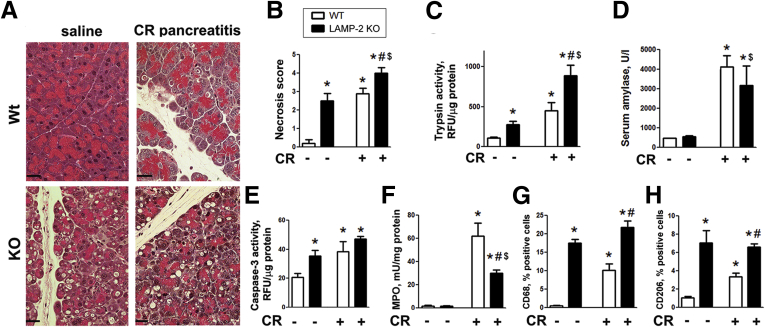
Results: Pancreatic levels of LAMP-1 and LAMP-2 greatly decrease across various pancreatitis models and in human disease. Pancreatitis does not trigger LAMPs' bulk de-glycosylation, but induces their degradation via CatB-mediated cleavage of LAMP molecule close to the boundary between luminal and transmembrane domains. LAMP-2 null mice spontaneously develop pancreatitis that begins with acinar cell vacuolization due to impaired autophagic flux, and progresses to severe pancreas damage characterized by trypsinogen activation, macrophage-driven inflammation, and acinar cell death. LAMP-2 deficiency causes a decrease in pancreatic digestive enzymes content, stimulates the basal and inhibits CCK-induced amylase secretion by acinar cells. The effects of LAMP-2 knockout and acute cerulein pancreatitis overlap, which corroborates the pathogenic role of LAMP decrease in experimental pancreatitis models.
Conclusions: The results indicate a critical role for LAMPs, particularly LAMP-2, in maintaining pancreatic acinar cell homeostasis, and provide evidence that defective lysosomal function, resulting in impaired autophagy, leads to pancreatitis. Mice with LAMP-2 deficiency present a novel genetic model of human pancreatitis caused by lysosomal/autophagic dysfunction.
Keywords: amylase secretion; autophagy; cathepsin B; cerulein.
Conflict of interest statement
: The authors disclose no conflicts
Figures
References
-
- Pandol S.J., Saluja A.K., Imrie C.W. Acute pancreatitis: bench to the bedside. Gastroenterology. 2007;132:1127–1151. [erratum 133:1056] - PubMed
-
- Aho H.J., Nevalainen T.J., Havia V.T. Human acute pancreatitis: a light and electron microscopic study. Acta Pathol Microbiol Immunol Scand A. 1982;90:367–373. - PubMed
-
- Helin H., Mero M., Markkula H. Pancreatic acinar ultrastructure in human acute pancreatitis. Virchows Arch A Pathol Anat Histol. 1980;387:259–270. - PubMed
-
- Watanabe O., Baccino F.M., Steer M.L. Supramaximal caerulein stimulation and ultrastructure of rat pancreatic acinar cell: early morphological changes during development of experimental pancreatitis. Am J Physiol. 1984;246:G457–G467. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous