

Chronic In Vivo Imaging of Ponto-Cerebellar Mossy Fibers Reveals Morphological Stability during Whisker Sensory Manipulation in the Adult Rat
- PMID: 26693178
- PMCID: PMC4676200
- DOI: 10.1523/ENEURO.0075-15.2015
Chronic In Vivo Imaging of Ponto-Cerebellar Mossy Fibers Reveals Morphological Stability during Whisker Sensory Manipulation in the Adult Rat
Abstract
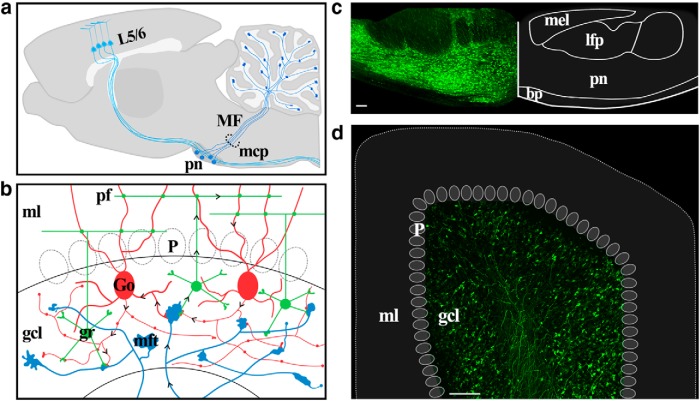
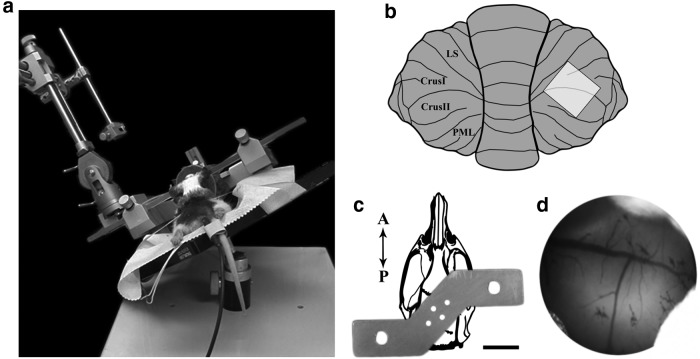
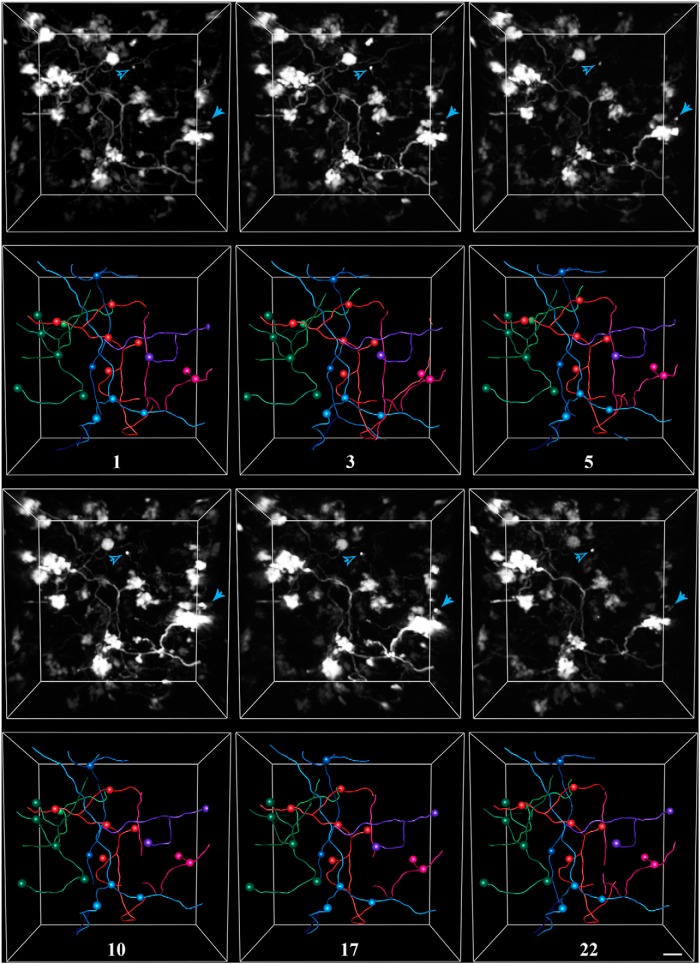
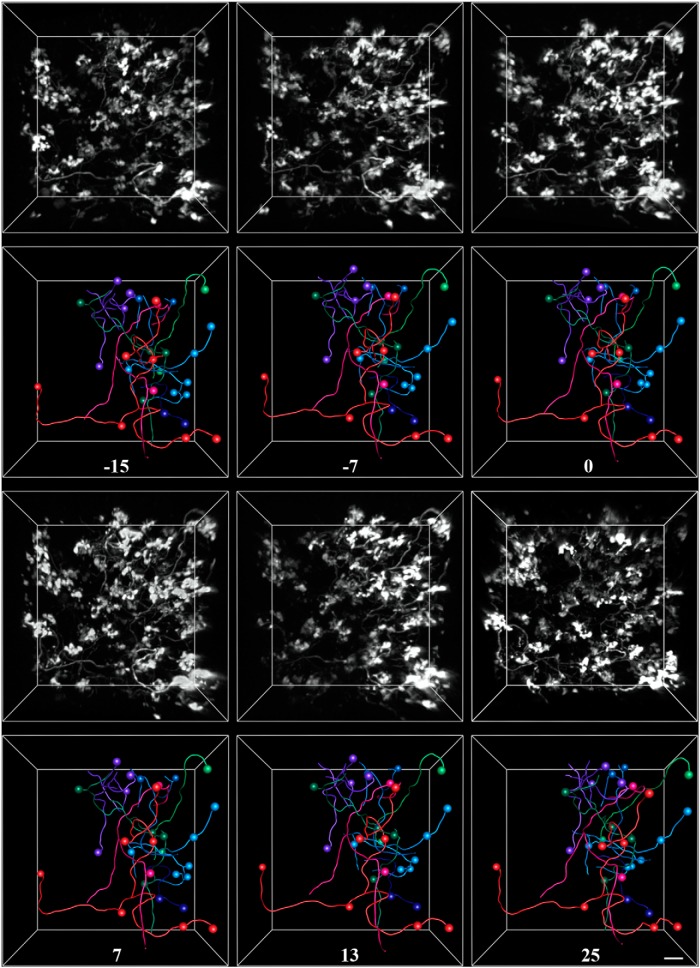
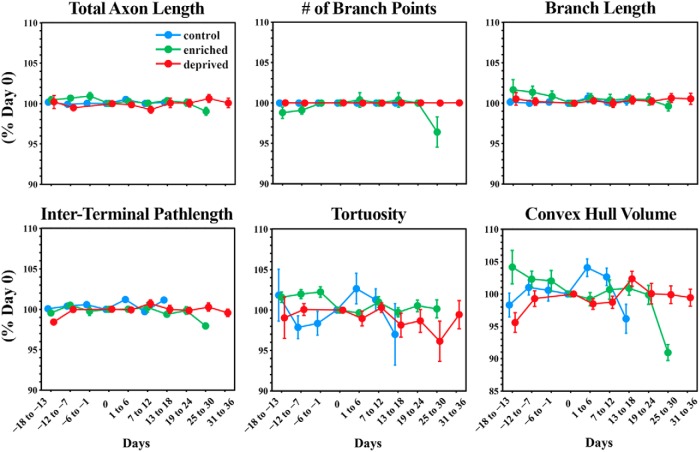
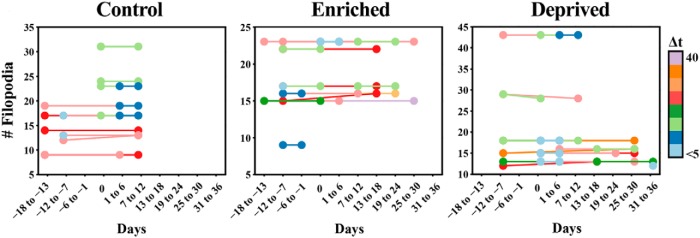
The cerebellum receives extensive disynaptic input from the neocortex via the basal pontine nuclei, the neurons of which send mossy fiber (MF) axons to the granule cell layer of the contralateral cerebellar hemisphere. Although this cortico-cerebellar circuit has been implicated in tasks such as sensory discrimination and motor learning, little is known about the potential role of MF morphological plasticity in the function of the cerebellar granule cell layer. To address this issue, we labeled MFs with EGFP via viral infection of the basal pons in adult rats and performed in vivo two-photon imaging of MFs in Crus I/II of the cerebellar hemisphere over a period of several weeks. Following the acquisition of baseline images, animals were housed in control, enriched, or deprived sensory environments. Morphological dynamics were assessed by tracing MF axons and their terminals, and by tracking the stability of filopodia arising from MF terminal rosettes. MF axons and terminals were found to be remarkably stable. Parameters derived neither from measurements of axonal arbor geometry nor from the morphology of individual rosettes and their filopodial extensions significantly changed under control conditions over 4 weeks of imaging. Increasing whisker stimulation by manipulating the sensory environment or decreasing such stimulation by whisker trimming also failed to alter MF structure. Our studies indicate that pontine MF axons projecting to Crus I/II in adult rats do not undergo significant structural rearrangements over the course of weeks, and that this stability is not altered by the sustained manipulation of whisker sensorimotor experience.
Keywords: cerebellum; in vivo imaging; mossy fiber.
Figures



Similar articles
-
Branching of individual somatosensory cerebropontine axons in rat: evidence of divergence.Brain Struct Funct. 2007 Jul;212(1):85-93. doi: 10.1007/s00429-007-0145-1. Epub 2007 May 25. Brain Struct Funct. 2007. PMID: 17717700
-
Axon collaterals of mossy fibers from the pontine nucleus in the cerebellar dentate nucleus.J Neurophysiol. 1992 Mar;67(3):547-60. doi: 10.1152/jn.1992.67.3.547. J Neurophysiol. 1992. PMID: 1578244
-
Divergent projections of single pontocerebellar axons to multiple cerebellar lobules in the mouse.J Comp Neurol. 2019 Aug 15;527(12):1966-1985. doi: 10.1002/cne.24662. Epub 2019 Feb 25. J Comp Neurol. 2019. PMID: 30737986
-
The Long Journey of Pontine Nuclei Neurons: From Rhombic Lip to Cortico-Ponto-Cerebellar Circuitry.Front Neural Circuits. 2017 May 17;11:33. doi: 10.3389/fncir.2017.00033. eCollection 2017. Front Neural Circuits. 2017. PMID: 28567005 Free PMC article. Review.
-
Mossy-fibre sensory input to the cerebellum.Prog Brain Res. 1997;114:251-9. doi: 10.1016/s0079-6123(08)63368-3. Prog Brain Res. 1997. PMID: 9193148 Review.
Cited by
-
The brain-specific RasGEF very-KIND is required for normal dendritic growth in cerebellar granule cells and proper motor coordination.PLoS One. 2017 Mar 6;12(3):e0173175. doi: 10.1371/journal.pone.0173175. eCollection 2017. PLoS One. 2017. PMID: 28264072 Free PMC article.
-
Developmental pattern and structural factors of dendritic survival in cerebellar granule cells in vivo.Sci Rep. 2018 Dec 3;8(1):17561. doi: 10.1038/s41598-018-35829-y. Sci Rep. 2018. PMID: 30510282 Free PMC article.
-
Spatiotemporal network coding of physiological mossy fiber inputs by the cerebellar granular layer.PLoS Comput Biol. 2017 Sep 21;13(9):e1005754. doi: 10.1371/journal.pcbi.1005754. eCollection 2017 Sep. PLoS Comput Biol. 2017. PMID: 28934196 Free PMC article.
-
Excitatory Cerebellar Nucleocortical Circuit Provides Internal Amplification during Associative Conditioning.Neuron. 2016 Feb 3;89(3):645-57. doi: 10.1016/j.neuron.2016.01.008. Neuron. 2016. PMID: 26844836 Free PMC article.
-
Optimal Degrees of Synaptic Connectivity.Neuron. 2017 Mar 8;93(5):1153-1164.e7. doi: 10.1016/j.neuron.2017.01.030. Epub 2017 Feb 16. Neuron. 2017. PMID: 28215558 Free PMC article.
References
-
- Allen GI, Tsukahara N (1974) Cerebrocerebellar communication systems. Physiol Rev 54:957-1006. - PubMed
-
- Beitz A, Larson A, Monaghan P, Altschuler R, Mullett M, Madl J (1986) Immunohistochemical localization of glutamate, glutaminase and aspartate-aminotransferase in neurons of the pontine nuclei of the rat. Neuroscience 17:741-753. - PubMed
-
- Bower JM, Beermann DH, Gibson JM, Shambes GM, Welker W (1981) Principles of organization of a cerebro-cerebellar circuit. micromapping the projections from cerebral (SI) to cerebellar (granule cell layer) tactile areas of rats. Brain Behav Evol 18:1-18. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources