

Dynamic spatio-temporal contribution of single β5t+ cortical epithelial precursors to the thymus medulla
- PMID: 26694097
- PMCID: PMC4832341
- DOI: 10.1002/eji.201545995
Dynamic spatio-temporal contribution of single β5t+ cortical epithelial precursors to the thymus medulla
Abstract
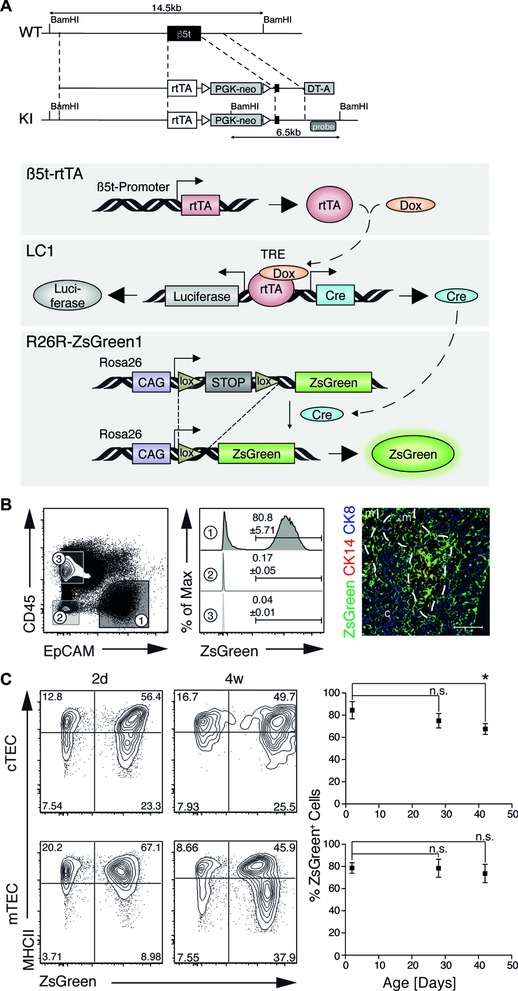
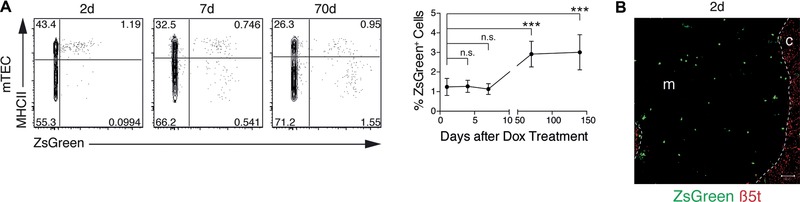
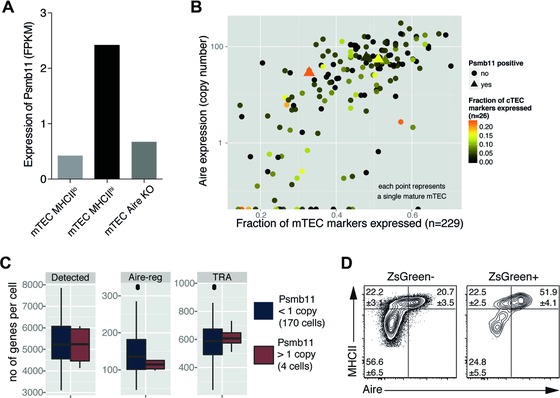
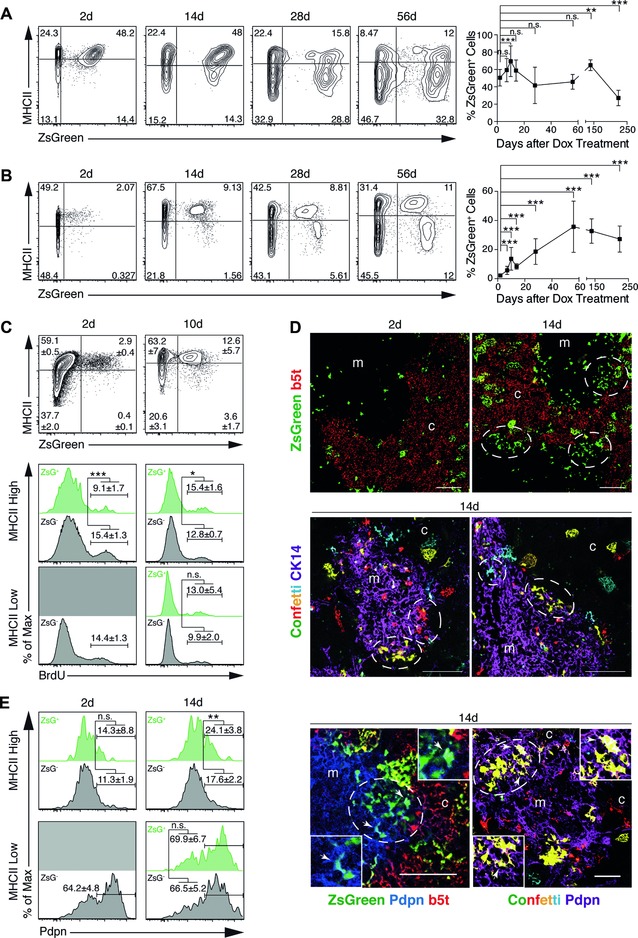
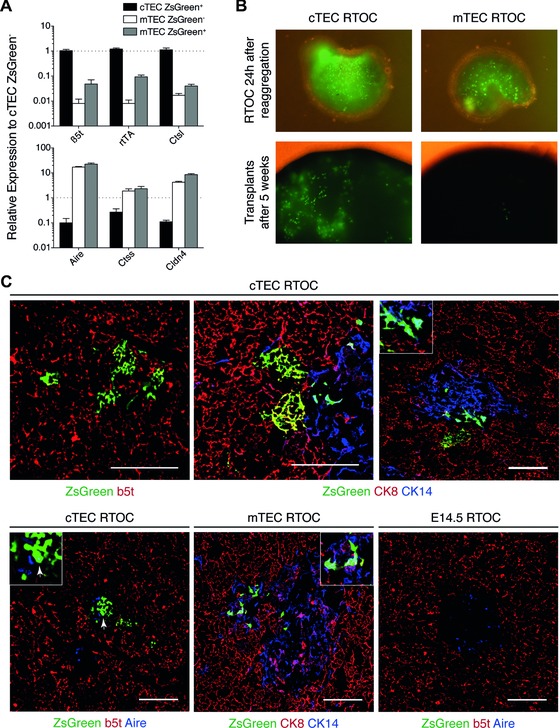
Intrathymic T-cell development is critically dependent on cortical and medullary thymic epithelial cells (TECs). Both epithelial subsets originate during early thymus organogenesis from progenitor cells that express the thymoproteasome subunit β5t, a typical feature of cortical TECs. Using in vivo lineage fate mapping, we demonstrate in mice that β5t(+) TEC progenitors give rise to the medullary TEC compartment early in life but significantly limit their contribution once the medulla has completely formed. Lineage-tracing studies at single cell resolution demonstrate for young mice that the postnatal medulla is expanded from individual β5t(+) cortical progenitors located at the cortico-medullary junction. These results therefore not only define a developmental window during which the expansion of medulla is efficiently enabled by progenitors resident in the thymic cortex, but also reveal the spatio-temporal dynamics that control the growth of the thymic medulla.
Keywords: Development; Epithelial cell; Medulla; Thymic progenitor cell; β5t.
© 2015 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
Comment in
-
Thymus medulla under construction: Time and space oddities.Eur J Immunol. 2016 Apr;46(4):829-33. doi: 10.1002/eji.201646329. Epub 2016 Mar 21. Eur J Immunol. 2016. PMID: 26947141
References
-
- Anderson, G. and Takahama, Y. , Thymic epithelial cells: working class heroes for T cell development and repertoire selection. Trends Immunol. 2012. 33: 256–263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
