

Rational design of crystal contact-free space in protein crystals for analyzing spatial distribution of motions within protein molecules
- PMID: 26694222
- PMCID: PMC4815401
- DOI: 10.1002/pro.2867
Rational design of crystal contact-free space in protein crystals for analyzing spatial distribution of motions within protein molecules
Abstract
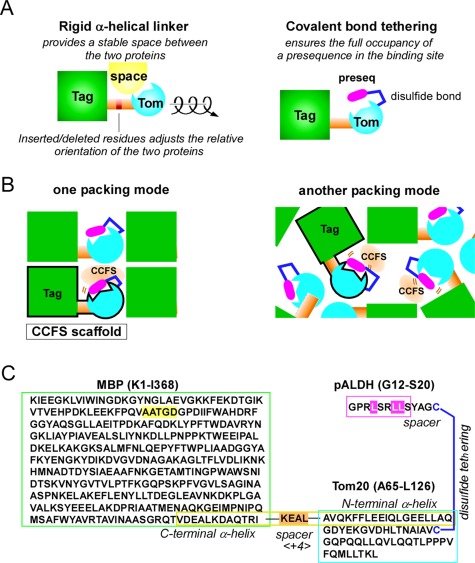
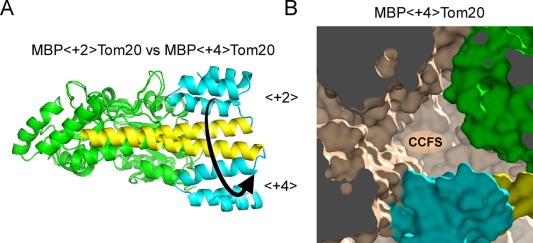
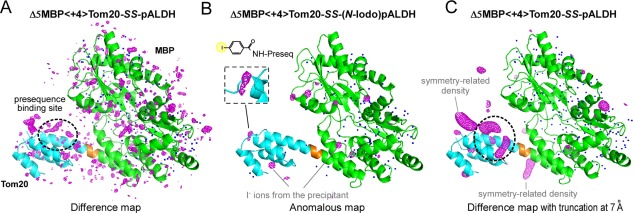
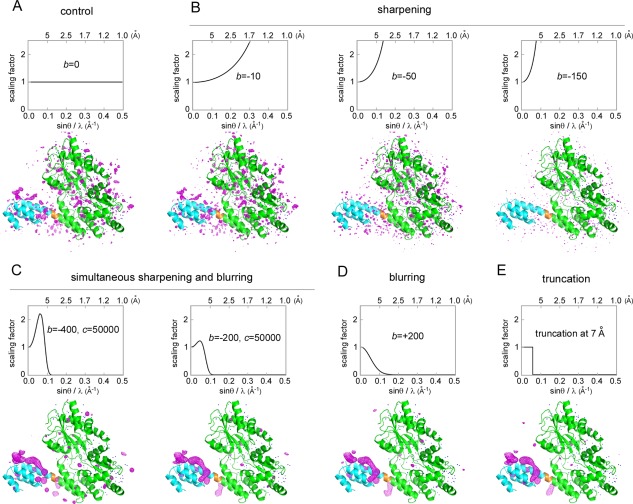
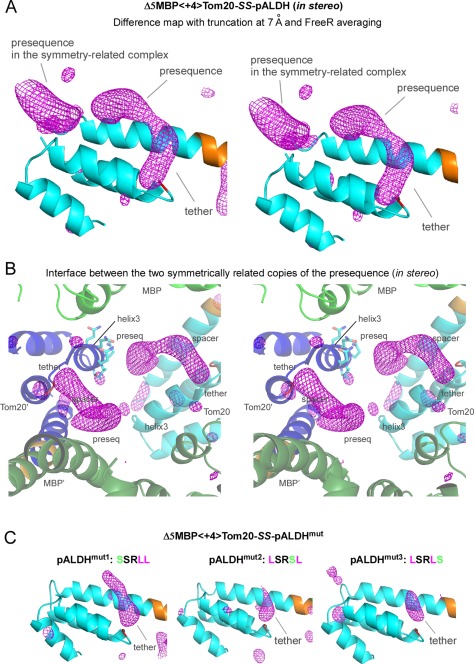
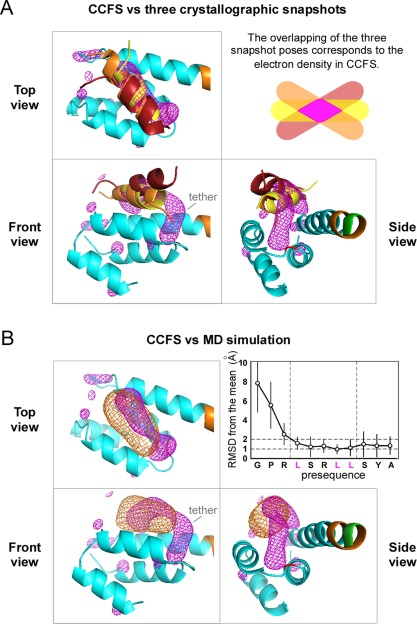
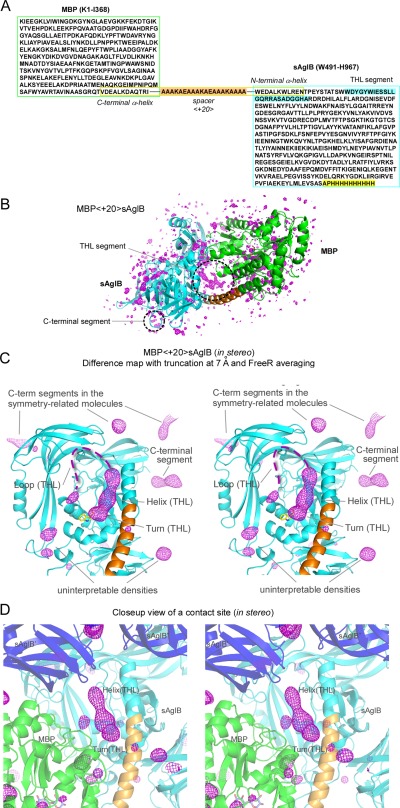
Contacts with neighboring molecules in protein crystals inevitably restrict the internal motions of intrinsically flexible proteins. The resultant clear electron densities permit model building, as crystallographic snapshot structures. Although these still images are informative, they could provide biased pictures of the protein motions. If the mobile parts are located at a site lacking direct contacts in rationally designed crystals, then the amplitude of the movements can be experimentally analyzed. We propose a fusion protein method, to create crystal contact-free space (CCFS) in protein crystals and to place the mobile parts in the CCFS. Conventional model building fails when large amplitude motions exist. In this study, the mobile parts appear as smeared electron densities in the CCFS, by suitable processing of the X-ray diffraction data. We applied the CCFS method to a highly mobile presequence peptide bound to the mitochondrial import receptor, Tom20, and a catalytically relevant flexible segment in the oligosaccharyltransferase, AglB. These two examples demonstrated the general applicability of the CCFS method to the analysis of the spatial distribution of motions within protein molecules.
Keywords: crystal contact effects; crystallographic method; fusion protein; mitochondrial presequence receptor Tom20; oligosaccharyltransferase AglB; protein motional analysis.
© 2016 The Protein Society.
Figures
References
-
- Ramanathan A, Savol A, Burger V, Chennubhotla CS, Agarwal PK (2014) Protein conformational populations and functionally relevant substates. Acc Chem Res 47:149–156. - PubMed
-
- Gobl C, Madl T, Simon B, Sattler M (2014) NMR approaches for structural analysis of multidomain proteins and complexes in solution. Prog Nucl Magn Reson Spectrosc 80:26–63. - PubMed
-
- Vranken WF (2014) NMR structure validation in relation to dynamics and structure determination. Prog Nucl Magn Reson Spectrosc 82:27–38. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
