

Beyond the Divide: Boundaries for Patterning and Stem Cell Regulation in Plants
- PMID: 26697027
- PMCID: PMC4673312
- DOI: 10.3389/fpls.2015.01052
Beyond the Divide: Boundaries for Patterning and Stem Cell Regulation in Plants
Abstract
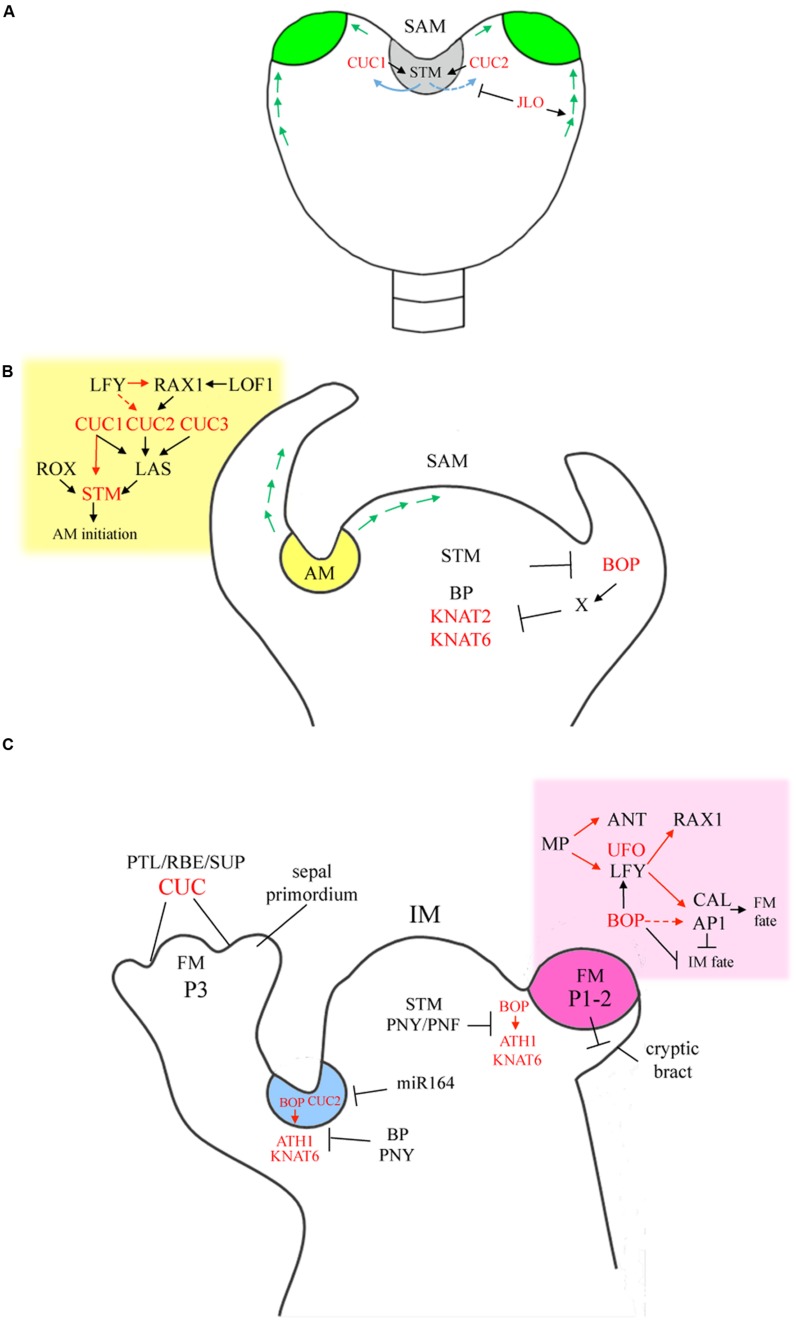
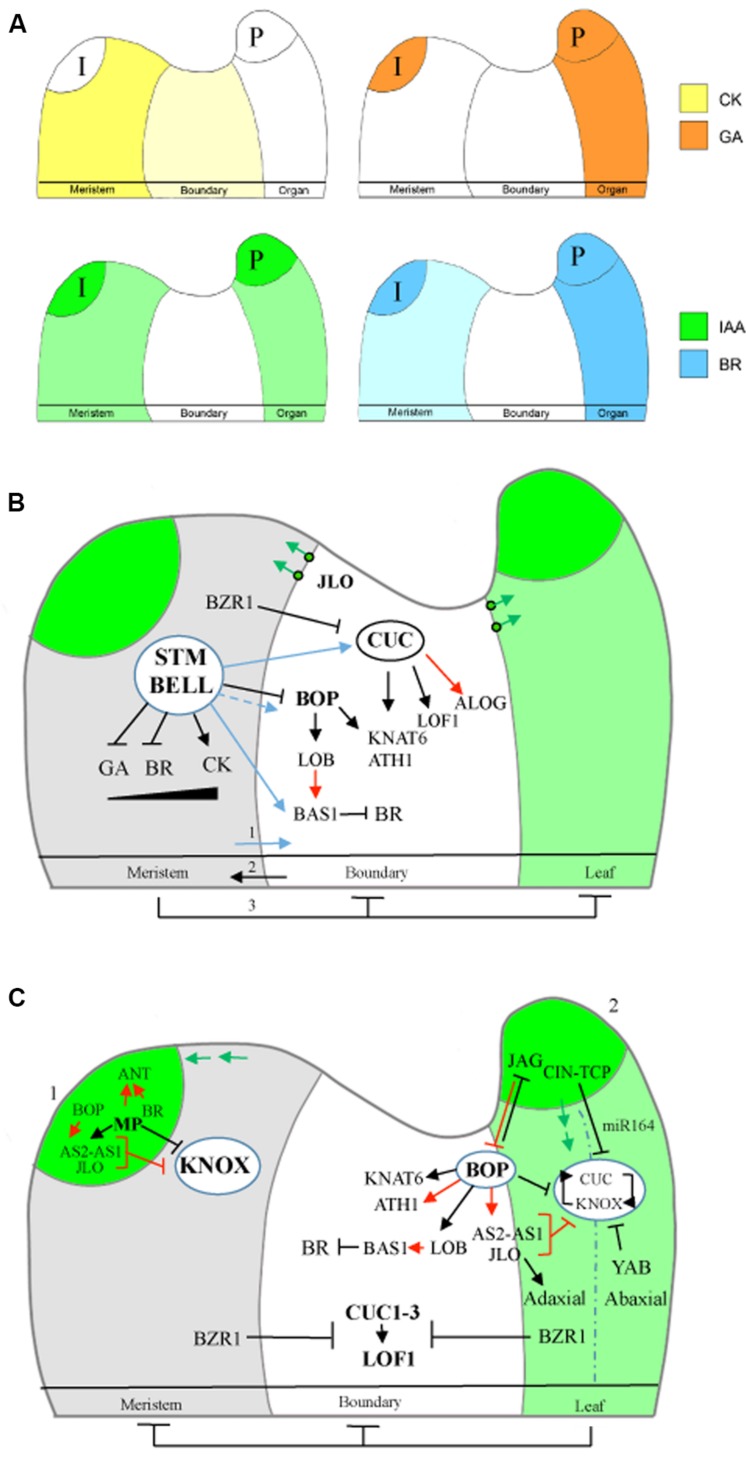
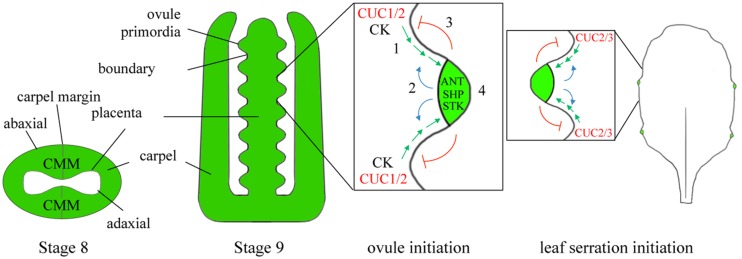
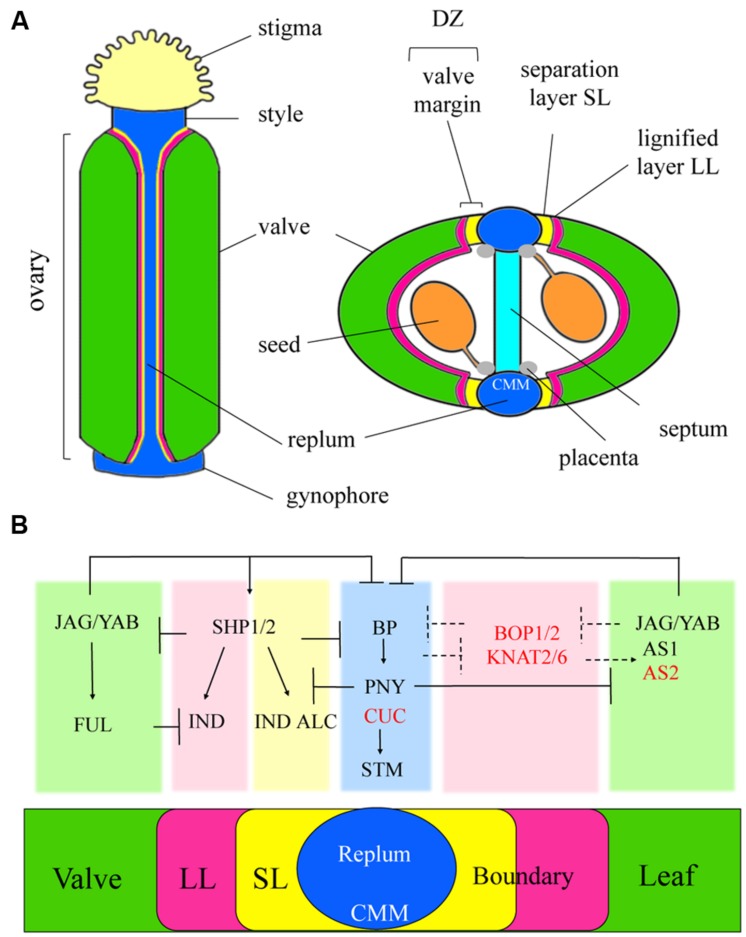
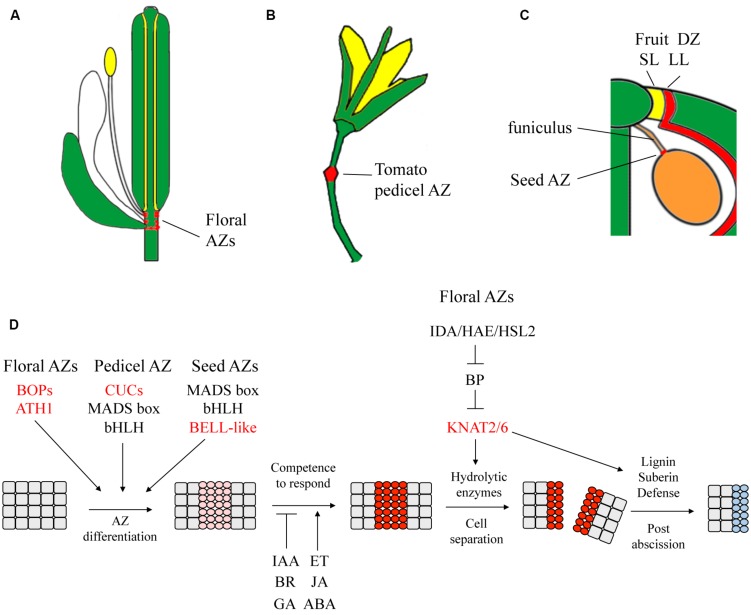
The initiation of plant lateral organs from the shoot apical meristem (SAM) is closely associated with the formation of specialized domains of restricted growth known as the boundaries. These zones are required in separating the meristem from the growing primordia or adjacent organs but play a much broader role in regulating stem cell activity and shoot patterning. Studies have revealed a network of genes and hormone pathways that establish and maintain boundaries between the SAM and leaves. Recruitment of these pathways is shown to underlie a variety of processes during the reproductive phase including axillary meristems production, flower patterning, fruit development, and organ abscission. This review summarizes the role of conserved gene modules in patterning boundaries throughout the life cycle.
Keywords: abscission; dehiscence; flower patterning; fruit patterning; inflorescence architecture; lateral organ boundary; meristem; organ separation.
Figures
References
-
- Aida M., Ishida T., Tasaka M. (1999). Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: interaction among the CUP-SHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development 126 1563–1570. - PubMed
-
- Aida M., Vernoux T., Furutani M., Traas J., Tasaka M. (2002). Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo. Development 129 3965–3974. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources