

Ectopic Expression in Arabidopsis thaliana of an NB-ARC Encoding Putative Disease Resistance Gene from Wild Chinese Vitis pseudoreticulata Enhances Resistance to Phytopathogenic Fungi and Bacteria
- PMID: 26697041
- PMCID: PMC4674559
- DOI: 10.3389/fpls.2015.01087
Ectopic Expression in Arabidopsis thaliana of an NB-ARC Encoding Putative Disease Resistance Gene from Wild Chinese Vitis pseudoreticulata Enhances Resistance to Phytopathogenic Fungi and Bacteria
Abstract
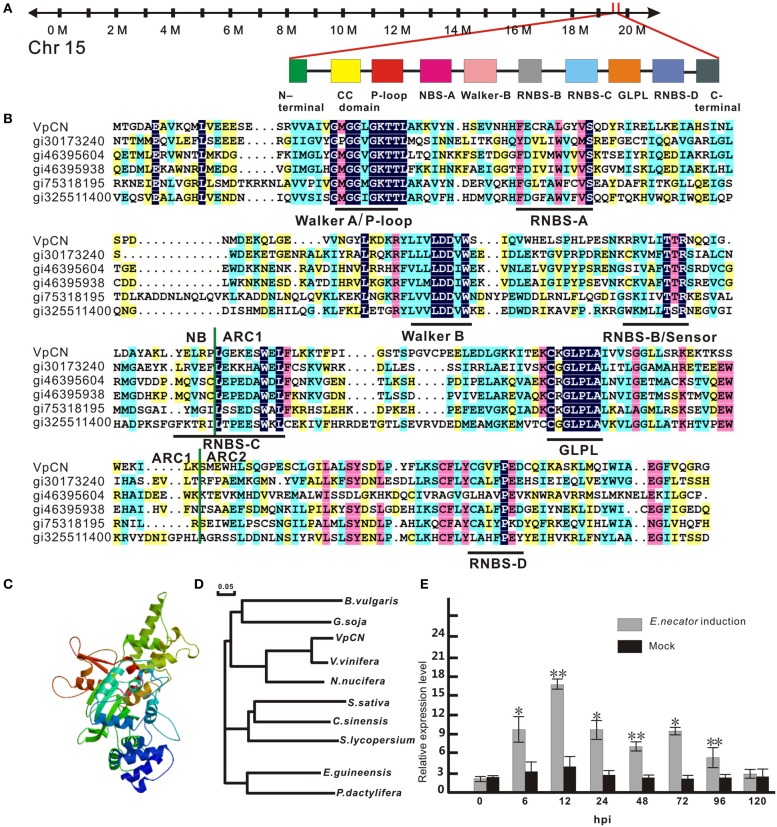
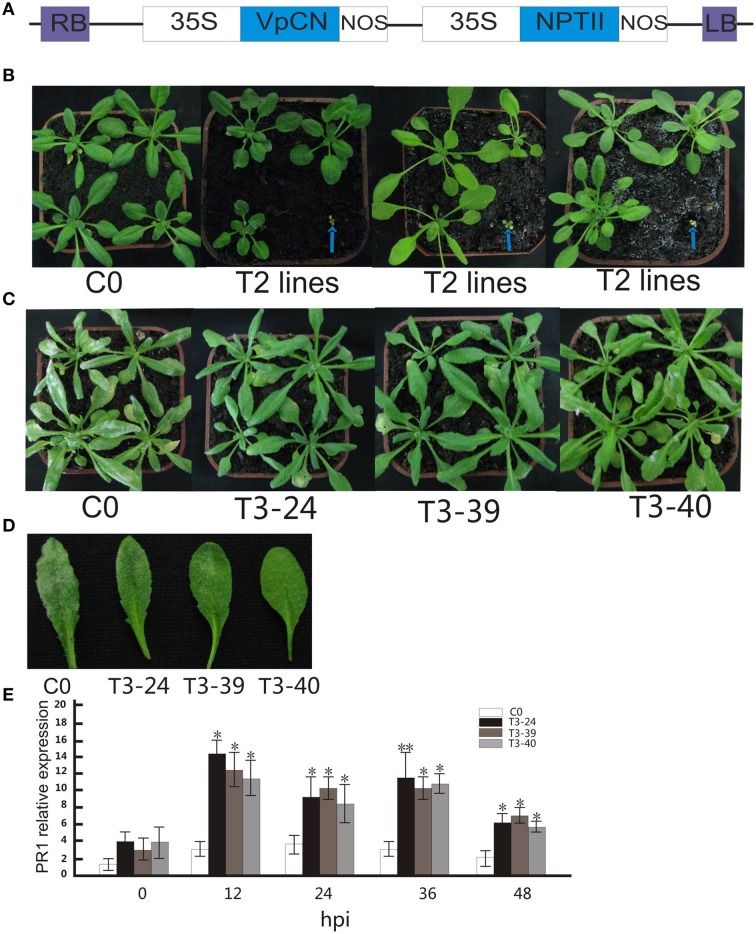
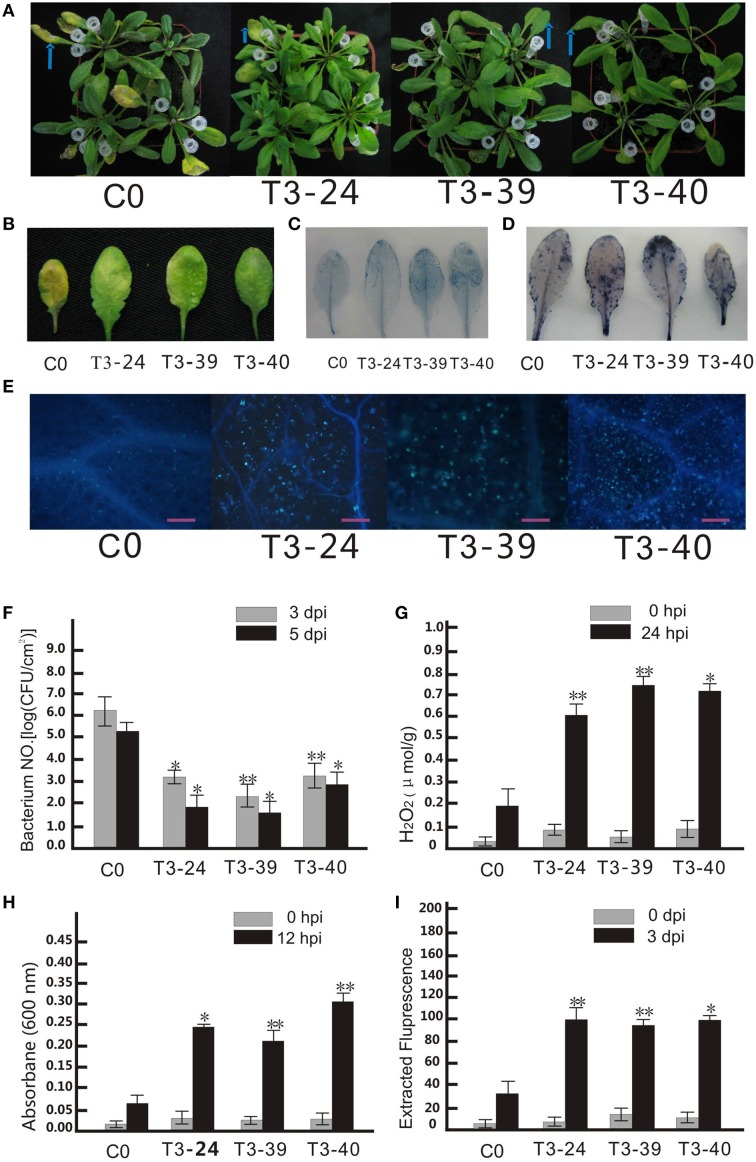
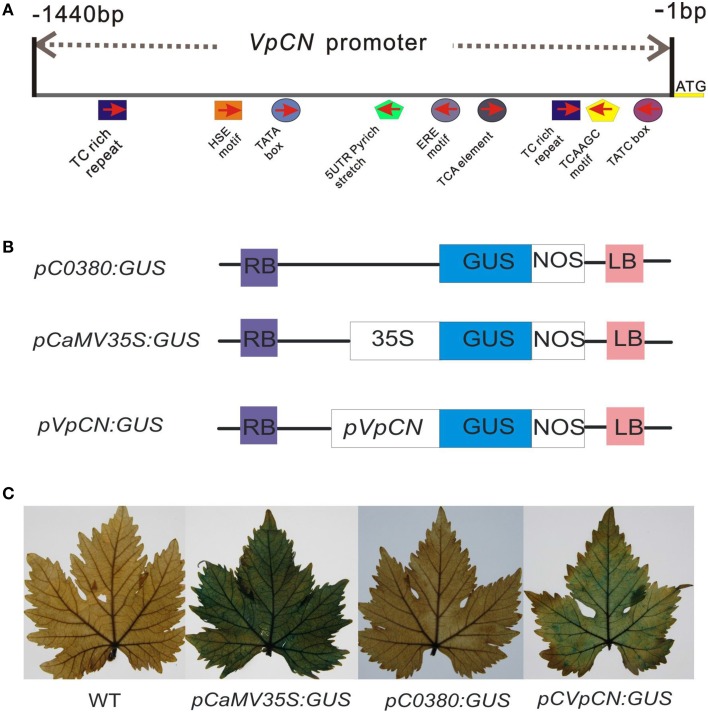
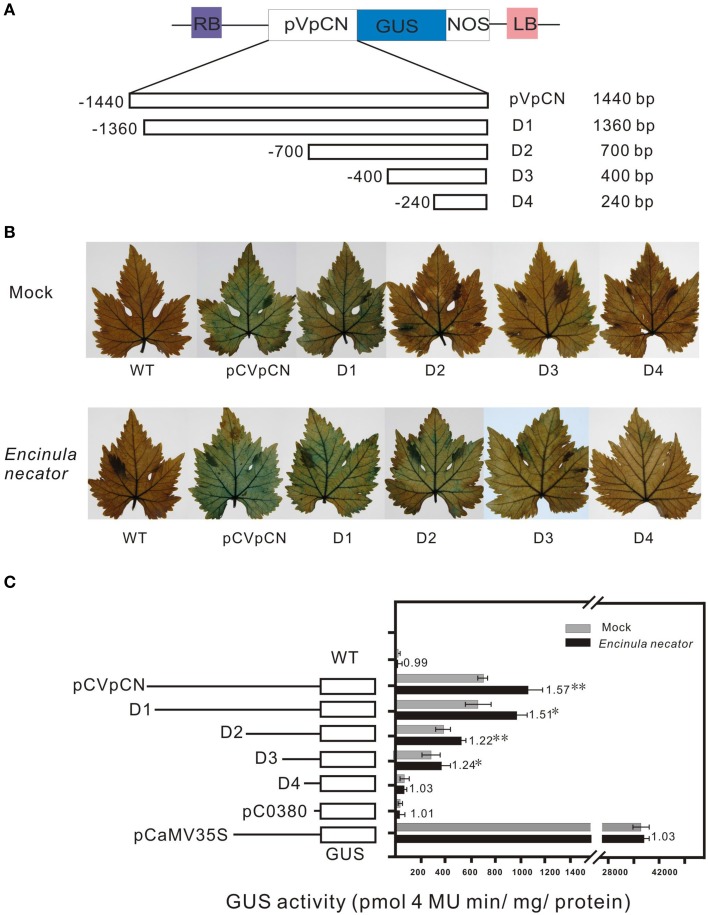
Plant resistance proteins mediate pathogen recognition and activate innate immune responses to restrict pathogen proliferation. One common feature of these proteins is an NB-ARC domain. In this study, we characterized a gene encoding a protein with an NB-ARC domain from wild Chinese grapevine Vitis pseudoreticulata accession "Baihe-35-1," which was identified in a transcriptome analysis of the leaves following inoculation with Erysiphe necator (Schw.), a causal agent of powdery mildew. Transcript levels of this gene, designated VpCN (GenBank accession number KT265084), increased strongly after challenge of grapevine leaves with E. necator. The deduced amino acid sequence was predicted to contain an NB-ARC domain in the C-terminus and an RxCC-like domain similar to CC domain of Rx protein in the N-terminus. Ectopic expression of VpCN in Arabidopsis thaliana resulted in either a wild-type phenotype or a dwarf phenotype. The phenotypically normal transgenic A. thaliana showed enhance resistance to A. thaliana powdery mildew Golovinomyces cichoracearum, as well as to a virulent bacterial pathogen Pseudomonas syringae pv. tomato DC3000. Moreover, promoter::GUS (β-glucuronidase) analysis revealed that powdery mildew infection induced the promoter activity of VpCN in grapevine leaves. Finally, a promoter deletion analysis showed that TC rich repeat elements likely play an important role in the response to E. necator infection. Taken together, our results suggest that VpCN contribute to powdery mildew disease resistant in grapevine.
Keywords: VpCN; disease resistance; powdery mildew; promoter analysis; wild Chinese Vitis.
Figures
Similar articles
-
Constitutive heterologous overexpression of a TIR-NB-ARC-LRR gene encoding a putative disease resistance protein from wild Chinese Vitis pseudoreticulata in Arabidopsis and tobacco enhances resistance to phytopathogenic fungi and bacteria.Plant Physiol Biochem. 2017 Mar;112:346-361. doi: 10.1016/j.plaphy.2017.01.017. Epub 2017 Jan 17. Plant Physiol Biochem. 2017. PMID: 28131063
-
VpRFP1, a novel C4C4-type RING finger protein gene from Chinese wild Vitis pseudoreticulata, functions as a transcriptional activator in defence response of grapevine.J Exp Bot. 2011 Nov;62(15):5671-82. doi: 10.1093/jxb/err253. Epub 2011 Aug 23. J Exp Bot. 2011. PMID: 21862480 Free PMC article.
-
The Chinese wild grapevine (Vitis pseudoreticulata) E3 ubiquitin ligase Erysiphe necator-induced RING finger protein 1 (EIRP1) activates plant defense responses by inducing proteolysis of the VpWRKY11 transcription factor.New Phytol. 2013 Nov;200(3):834-846. doi: 10.1111/nph.12418. Epub 2013 Jul 31. New Phytol. 2013. PMID: 23905547
-
Powdery Mildew Resistance Genes in Vines: An Opportunity to Achieve a More Sustainable Viticulture.Pathogens. 2022 Jun 18;11(6):703. doi: 10.3390/pathogens11060703. Pathogens. 2022. PMID: 35745557 Free PMC article. Review.
-
Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease.Hortic Res. 2015 May 20;2:15020. doi: 10.1038/hortres.2015.20. eCollection 2015. Hortic Res. 2015. PMID: 26504571 Free PMC article. Review.
Cited by
-
Comparative transcriptome analysis reveals whole-genome duplications and gene selection patterns in cultivated and wild Chrysanthemum species.Plant Mol Biol. 2017 Nov;95(4-5):451-461. doi: 10.1007/s11103-017-0663-z. Epub 2017 Oct 19. Plant Mol Biol. 2017. PMID: 29052098 Free PMC article.
-
Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.).Plants (Basel). 2023 Dec 29;13(1):110. doi: 10.3390/plants13010110. Plants (Basel). 2023. PMID: 38202418 Free PMC article.
-
Deep sequencing and in silico analysis of small RNA library reveals novel miRNA from leaf Persicaria minor transcriptome.3 Biotech. 2018 Mar;8(3):136. doi: 10.1007/s13205-018-1164-8. Epub 2018 Feb 15. 3 Biotech. 2018. PMID: 29479512 Free PMC article.
-
Molecular cloning of a CC-NBS-LRR gene from Vitis quinquangularis and its expression pattern in response to downy mildew pathogen infection.Mol Genet Genomics. 2018 Feb;293(1):61-68. doi: 10.1007/s00438-017-1360-y. Epub 2017 Sep 1. Mol Genet Genomics. 2018. PMID: 28864888
-
Emergent Ascomycetes in Viticulture: An Interdisciplinary Overview.Front Plant Sci. 2019 Nov 22;10:1394. doi: 10.3389/fpls.2019.01394. eCollection 2019. Front Plant Sci. 2019. PMID: 31824521 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources