

Gamma-actin is involved in regulating centrosome function and mitotic progression in cancer cells
- PMID: 26697841
- PMCID: PMC4825712
- DOI: 10.1080/15384101.2015.1120920
Gamma-actin is involved in regulating centrosome function and mitotic progression in cancer cells
Abstract
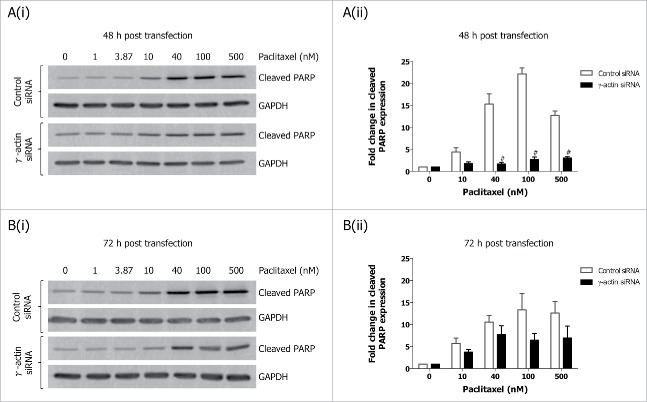
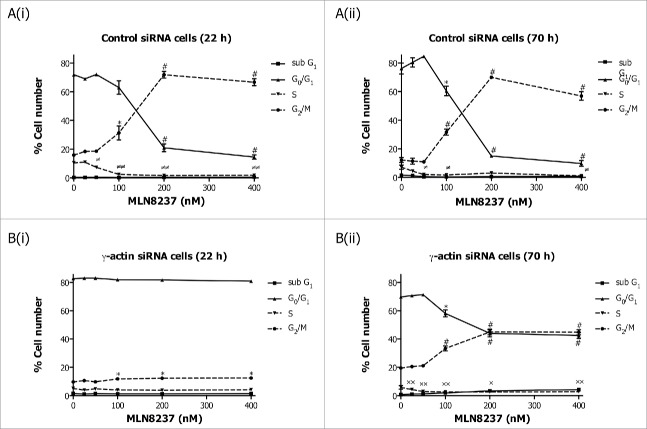
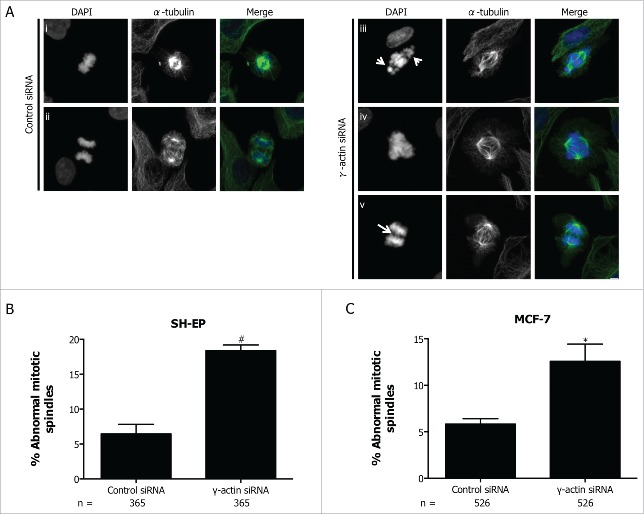
Reorganization of the actin cytoskeleton during mitosis is crucial for regulating cell division. A functional role for γ-actin in mitotic arrest induced by the microtubule-targeted agent, paclitaxel, has recently been demonstrated. We hypothesized that γ-actin plays a role in mitosis. Herein, we investigated the effect of γ-actin in mitosis and demonstrated that γ-actin is important in the distribution of β-actin and formation of actin-rich retraction fibers during mitosis. The reduced ability of paclitaxel to induce mitotic arrest as a result of γ-actin depletion was replicated with a range of mitotic inhibitors, suggesting that γ-actin loss reduces the ability of broad classes of anti-mitotic agents to induce mitotic arrest. In addition, partial depletion of γ-actin enhanced centrosome amplification in cancer cells and caused a significant delay in prometaphase/metaphase. This prolonged prometaphase/metaphase arrest was due to mitotic defects such as uncongressed and missegregated chromosomes, and correlated with an increased presence of mitotic spindle abnormalities in the γ-actin depleted cells. Collectively, these results demonstrate a previously unknown role for γ-actin in regulating centrosome function, chromosome alignment and maintenance of mitotic spindle integrity.
Keywords: abnormal mitotic spindles; centrosome amplification; mitosis; mitotic abnormalities; mitotic progression; γ-actin.
Figures





Comment in
-
Revisiting Actin's role in early centrosome separation.Cell Cycle. 2016;15(2):162-3. doi: 10.1080/15384101.2015.1130090. Cell Cycle. 2016. PMID: 26701337 Free PMC article. No abstract available.
References
-
- Heng YW, Koh CG. Actin cytoskeleton dynamics and the cell division cycle. Int J Biochem Cell Biol 2010; 42:1622-33; PMID:20412868; http://dx.doi.org/ 10.1016/j.biocel.2010.04.007 - DOI - PubMed
-
- Rankin KE, Wordeman L. Long astral microtubules uncouple mitotic spindles from the cytokinetic furrow. J Cell Biol 2010; 190:35-43; PMID:20603328; http://dx.doi.org/ 10.1083/jcb.201004017 - DOI - PMC - PubMed
-
- Palmer RE, Sullivan DS, Huffaker T, Koshland D. Role of astral microtubules and actin in spindle orientation and migration in the budding yeast, Saccharomyces cerevisiae. J Cell Biol 1992; 119:583-93; PMID:1400594; http://dx.doi.org/ 10.1083/jcb.119.3.583 - DOI - PMC - PubMed
-
- Sandquist JC, Kita AM, Bement WM. And the dead shall rise: actin and myosin return to the spindle. Dev Cell 2011; 21:410-9; PMID:21920311; http://dx.doi.org/ 10.1016/j.devcel.2011.07.018 - DOI - PMC - PubMed
-
- Weber KL, Sokac AM, Berg JS, Cheney RE, Bement WM. A microtubule-binding myosin required for nuclear anchoring and spindle assembly. Nature 2004; 431:325-9; PMID:15372037; http://dx.doi.org/ 10.1038/nature02834 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources