

Autophagy in the CNS and Periphery Coordinate Lipophagy and Lipolysis in the Brown Adipose Tissue and Liver
- PMID: 26698918
- PMCID: PMC4715637
- DOI: 10.1016/j.cmet.2015.10.008
Autophagy in the CNS and Periphery Coordinate Lipophagy and Lipolysis in the Brown Adipose Tissue and Liver
Abstract
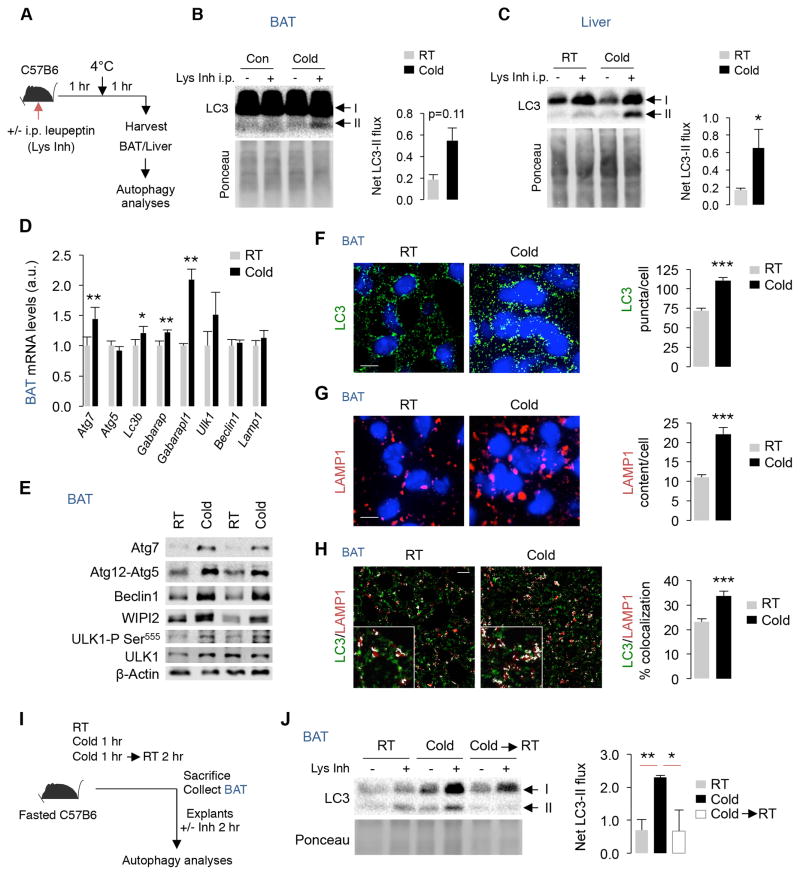
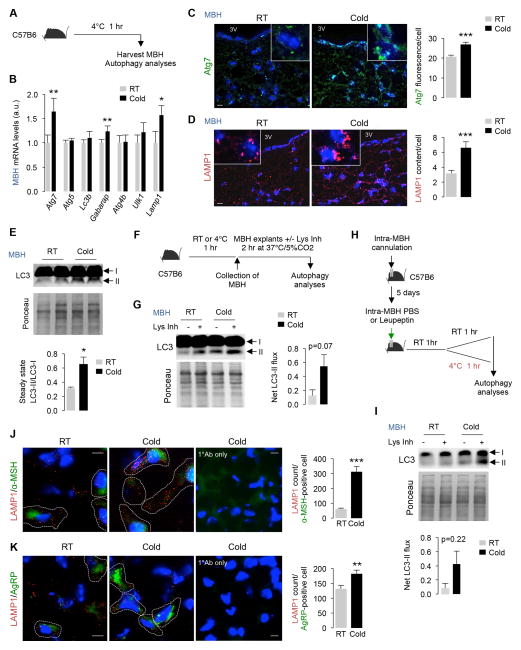
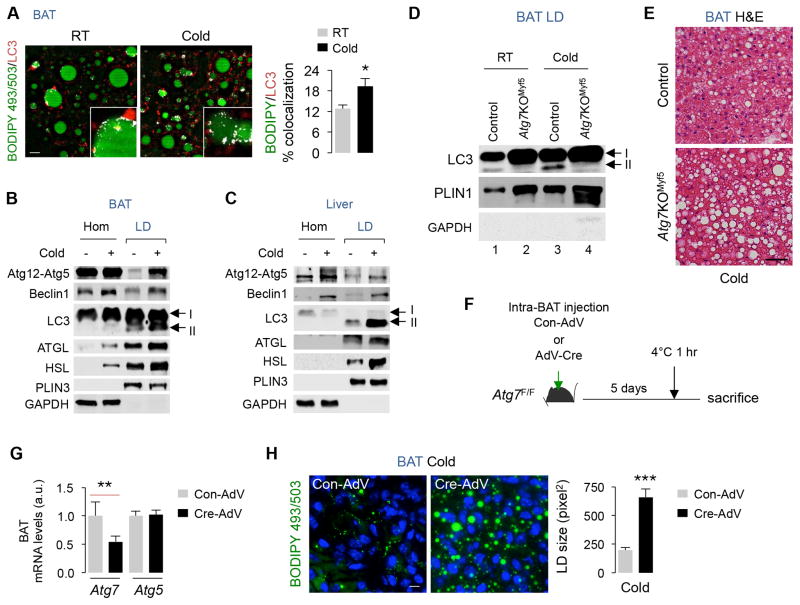
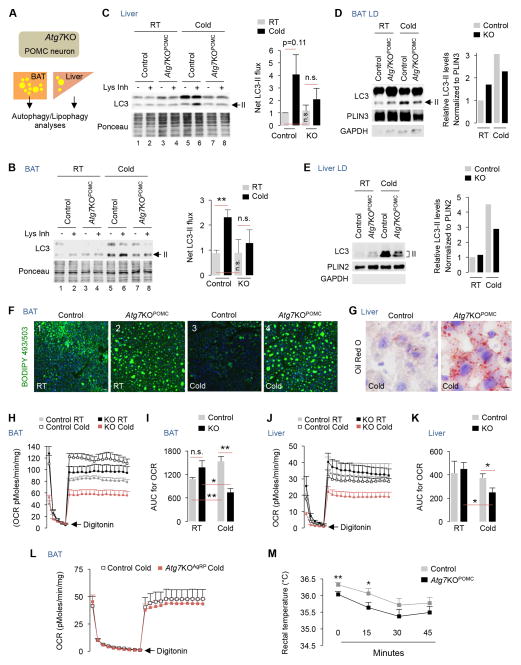
The integrative physiology of inter-organ communication in lipophagy regulation is not well understood. Lipophagy and the cytosolic lipases ATGL and HSL contribute to lipid droplet (LD) mobilization; however, whether autophagy proteins engage with lipases to promote lipid utilization remains unknown. Here, we show that cold induces autophagy in proopiomelanocortin (POMC) neurons and activates lipophagy in brown adipose tissue (BAT) and liver in mice. Targeted activation of autophagy in POMC neurons via intra-hypothalamic rapamycin is sufficient to trigger lipid utilization in room temperature-housed mice. Conversely, inhibiting autophagy in POMC neurons or in peripheral tissues or denervating BAT blocks lipid utilization. Unexpectedly, the autophagosome marker LC3 is mechanistically coupled to ATGL-mediated lipolysis. ATGL exhibits LC3-interacting region (LIR) motifs, and mutating a single LIR motif on ATGL displaces ATGL from LD and disrupts lipolysis. Thus, cold-induced activation of central autophagy activates lipophagy and cytosolic lipases in a complementary manner to mediate lipolysis in peripheral tissues.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors have no competing financial interests to declare.
Figures
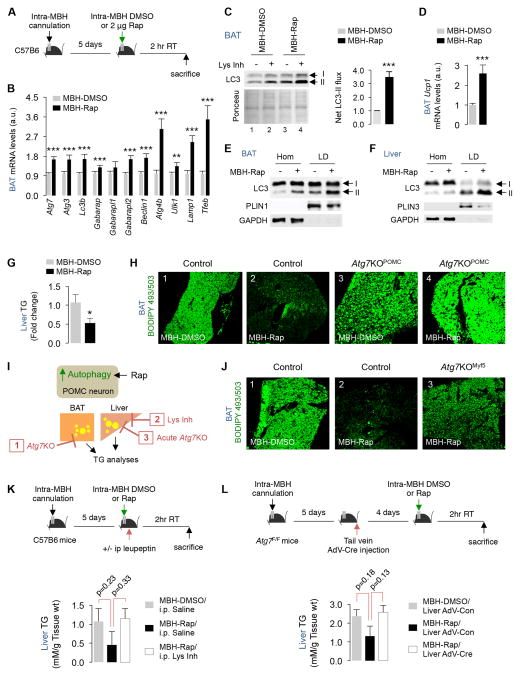
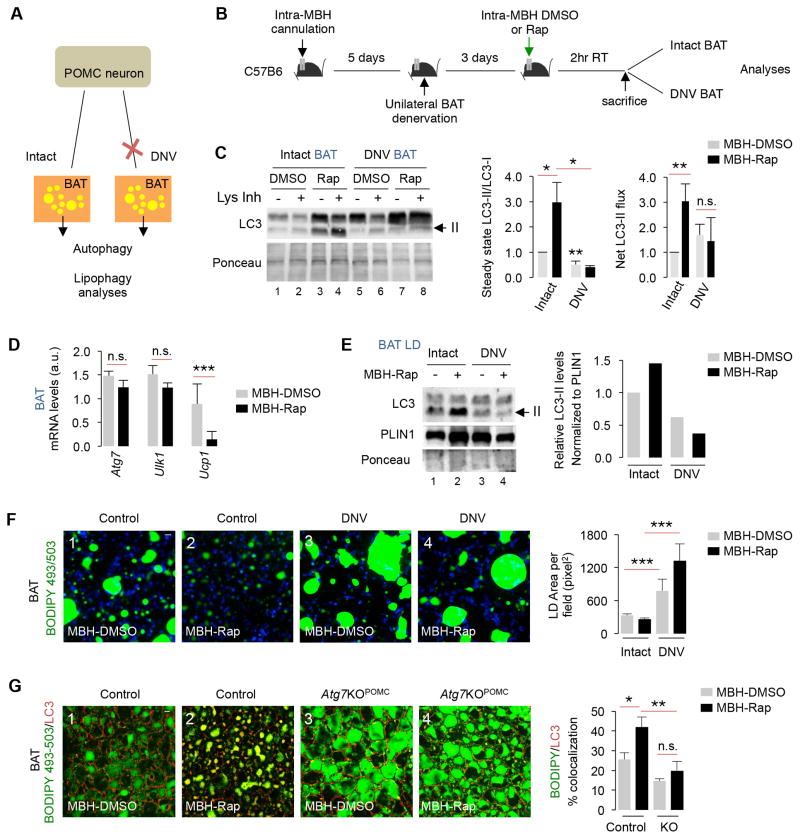
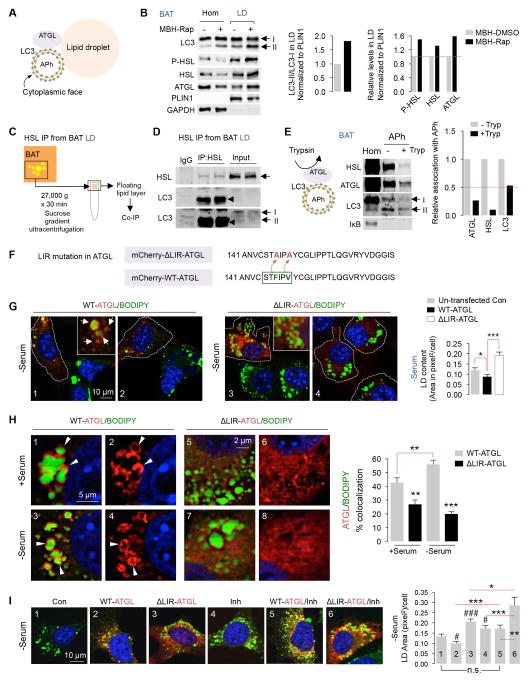
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
