

Nicastrin functions to sterically hinder γ-secretase-substrate interactions driven by substrate transmembrane domain
- PMID: 26699478
- PMCID: PMC4747693
- DOI: 10.1073/pnas.1512952113
Nicastrin functions to sterically hinder γ-secretase-substrate interactions driven by substrate transmembrane domain
Abstract
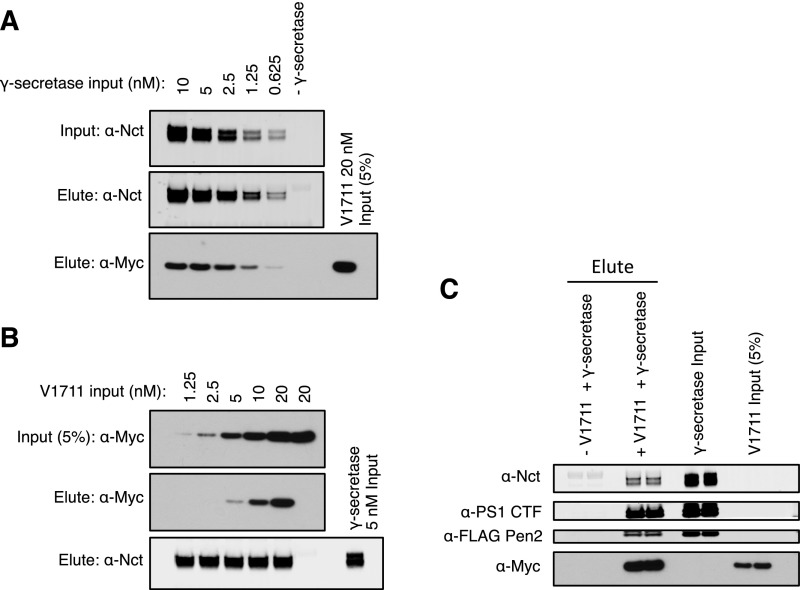
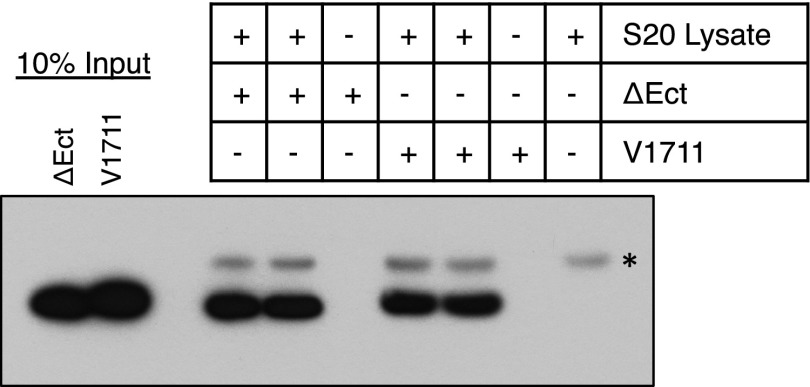
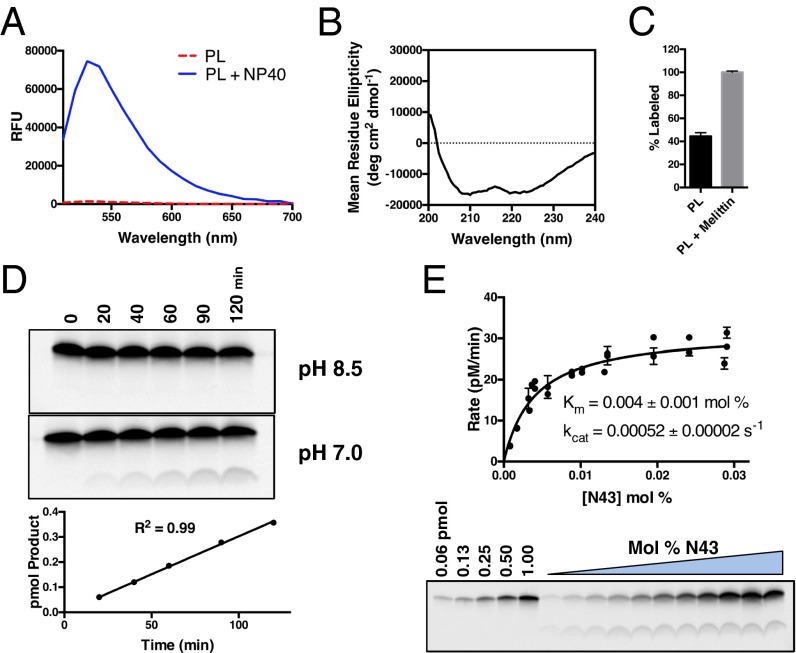
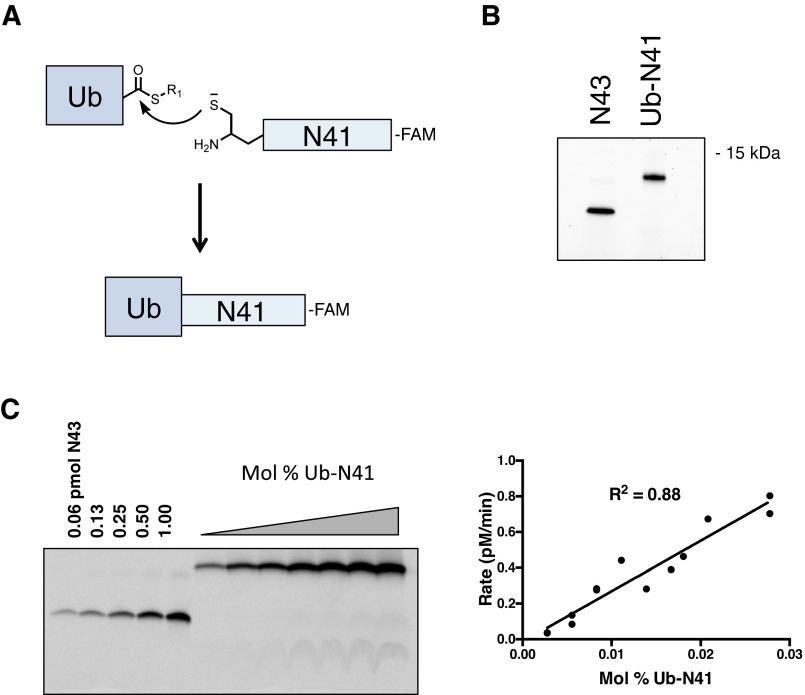
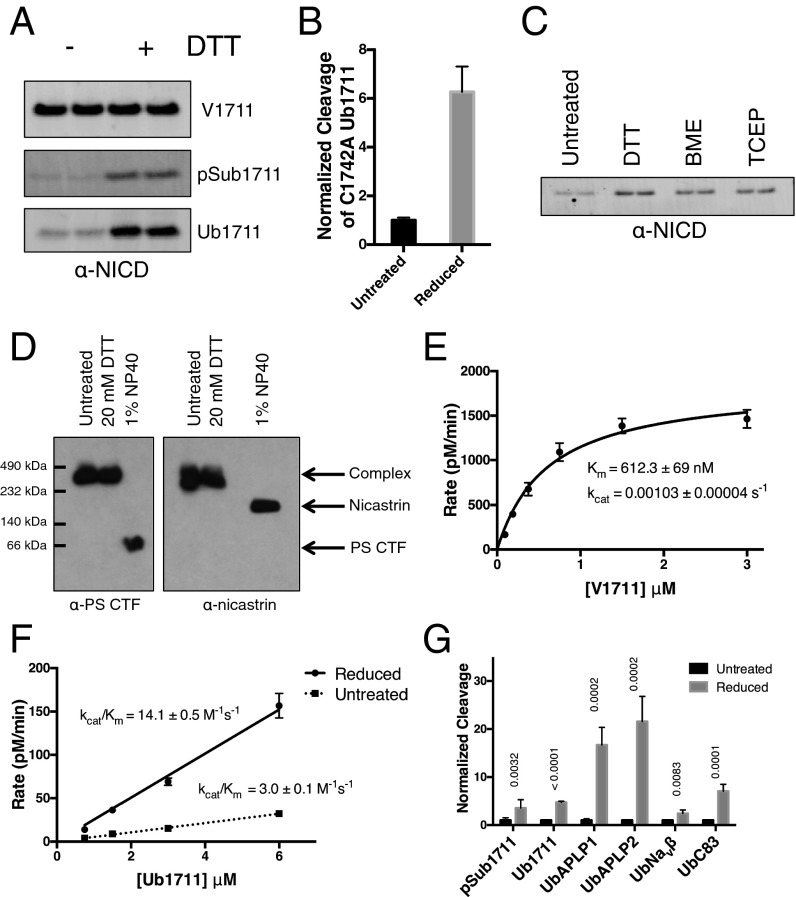
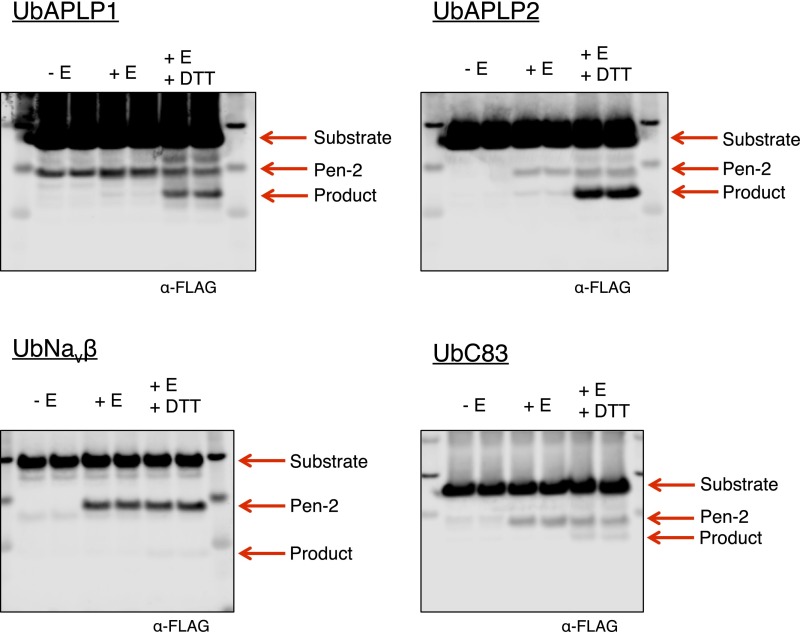
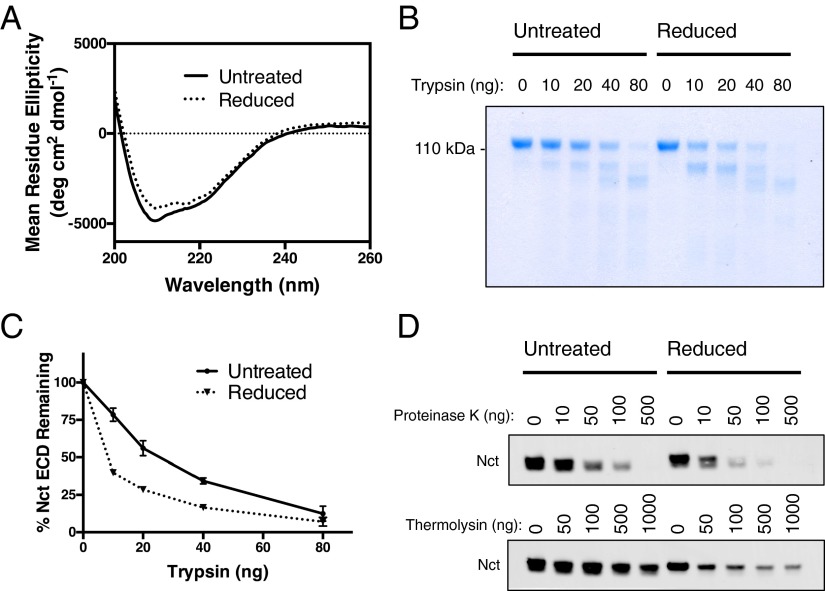
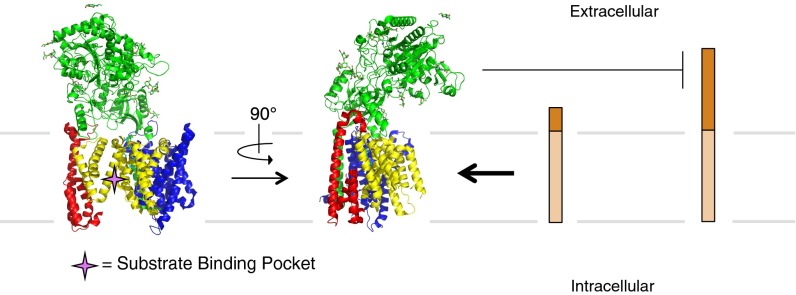
γ-Secretase is an intramembrane-cleaving protease that processes many type-I integral membrane proteins within the lipid bilayer, an event preceded by shedding of most of the substrate's ectodomain by α- or β-secretases. The mechanism by which γ-secretase selectively recognizes and recruits ectodomain-shed substrates for catalysis remains unclear. In contrast to previous reports that substrate is actively recruited for catalysis when its remaining short ectodomain interacts with the nicastrin component of γ-secretase, we find that substrate ectodomain is entirely dispensable for cleavage. Instead, γ-secretase-substrate binding is driven by an apparent tight-binding interaction derived from substrate transmembrane domain, a mechanism in stark contrast to rhomboid--another family of intramembrane-cleaving proteases. Disruption of the nicastrin fold allows for more efficient cleavage of substrates retaining longer ectodomains, indicating that nicastrin actively excludes larger substrates through steric hindrance, thus serving as a molecular gatekeeper for substrate binding and catalysis.
Keywords: Azheimer’s disease; intramembrane-cleaving protease; nicastrin; notch; γ-secretase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
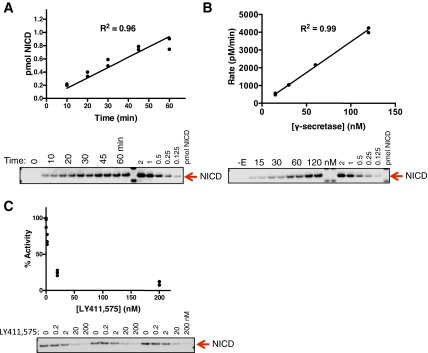
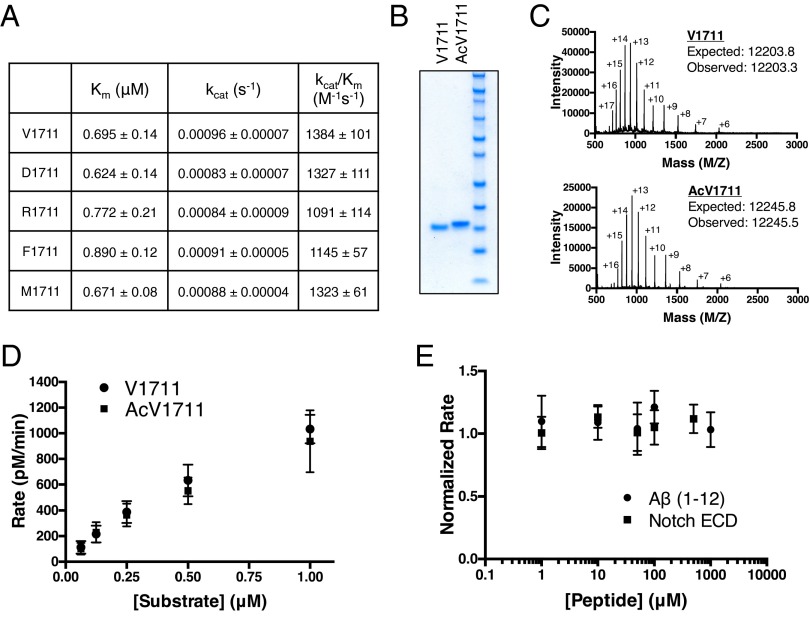
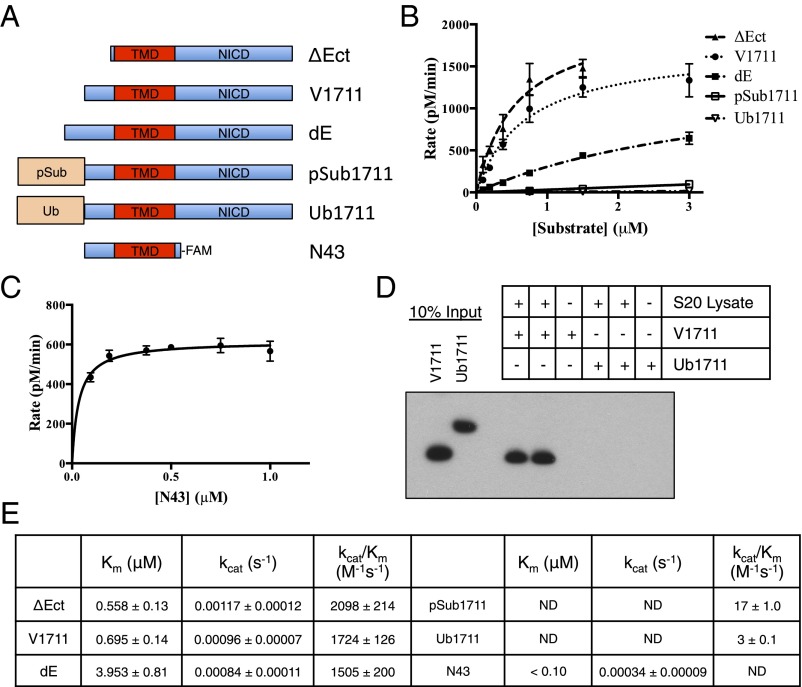
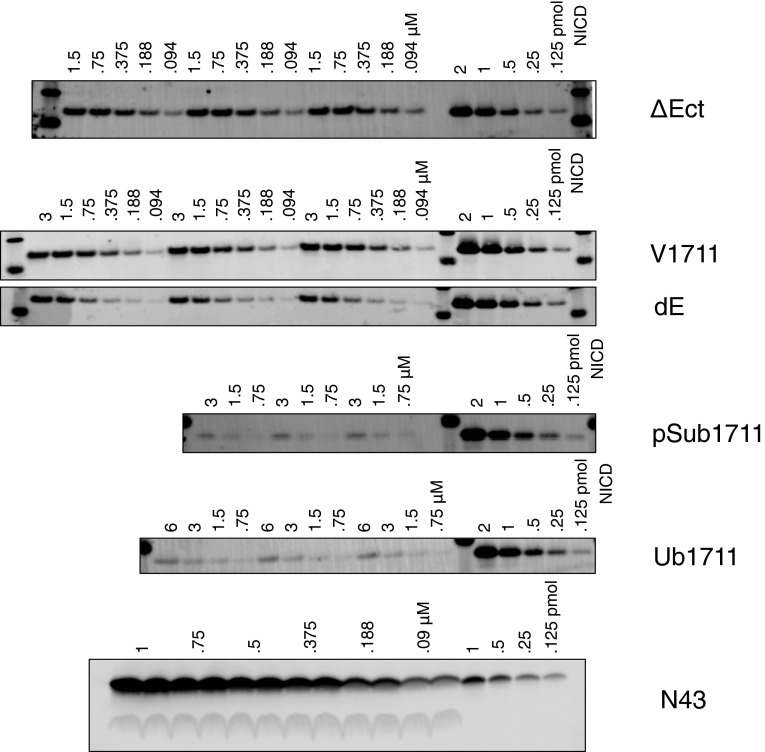

Comment in
-
Nicastrin guards Alzheimer's gate.Proc Natl Acad Sci U S A. 2016 Feb 2;113(5):1112-4. doi: 10.1073/pnas.1524151113. Epub 2016 Jan 25. Proc Natl Acad Sci U S A. 2016. PMID: 26811479 Free PMC article. No abstract available.
References
-
- Urban S. Mechanisms and cellular functions of intramembrane proteases. Biochim Biophys Acta. 2013;1828(12):2797–2800. - PubMed
-
- Huppert SS, et al. Embryonic lethality in mice homozygous for a processing-deficient allele of Notch1. Nature. 2000;405(6789):966–970. - PubMed
-
- Rawson RB, et al. Complementation cloning of S2P, a gene encoding a putative metalloprotease required for intramembrane cleavage of SREBPs. Mol Cell. 1997;1(1):47–57. - PubMed
-
- Ye J, et al. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol Cell. 2000;6(6):1355–1364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
