

Effects of aging on circadian patterns of gene expression in the human prefrontal cortex
- PMID: 26699485
- PMCID: PMC4711850
- DOI: 10.1073/pnas.1508249112
Effects of aging on circadian patterns of gene expression in the human prefrontal cortex
Abstract
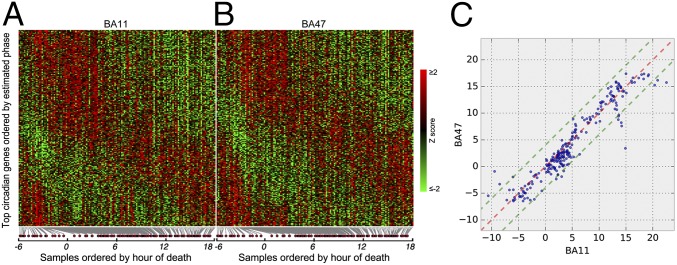
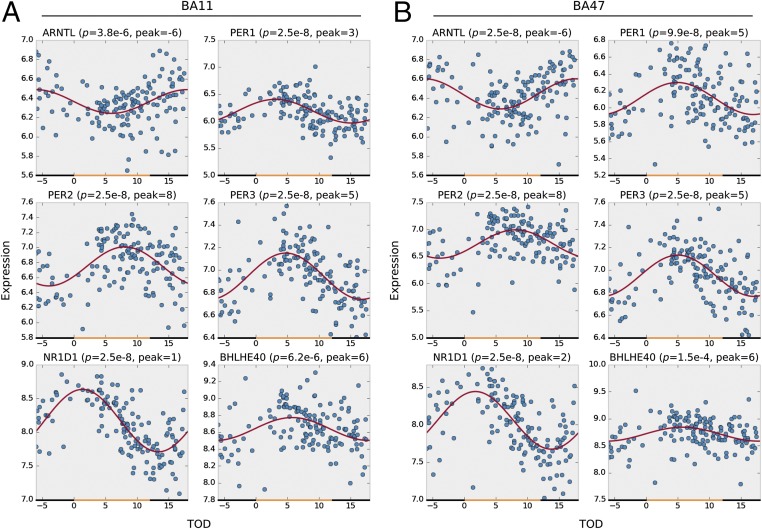
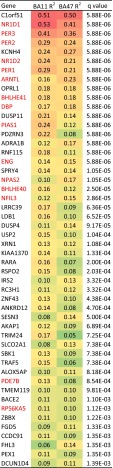
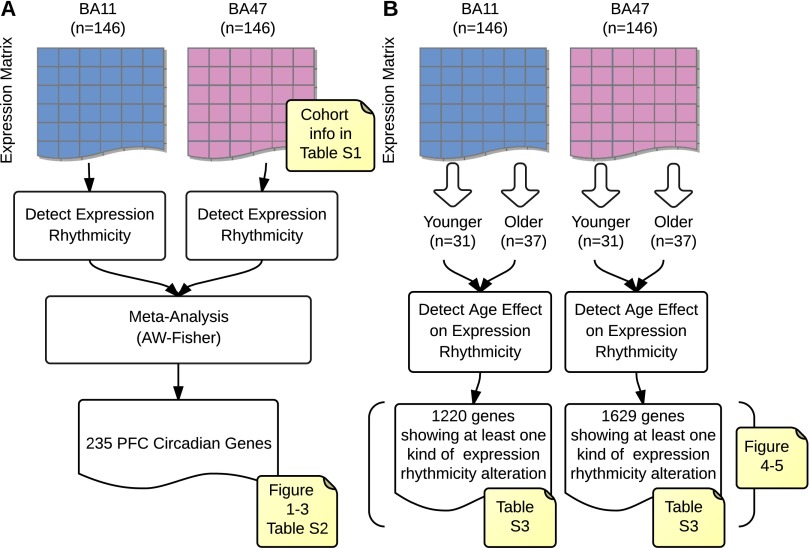
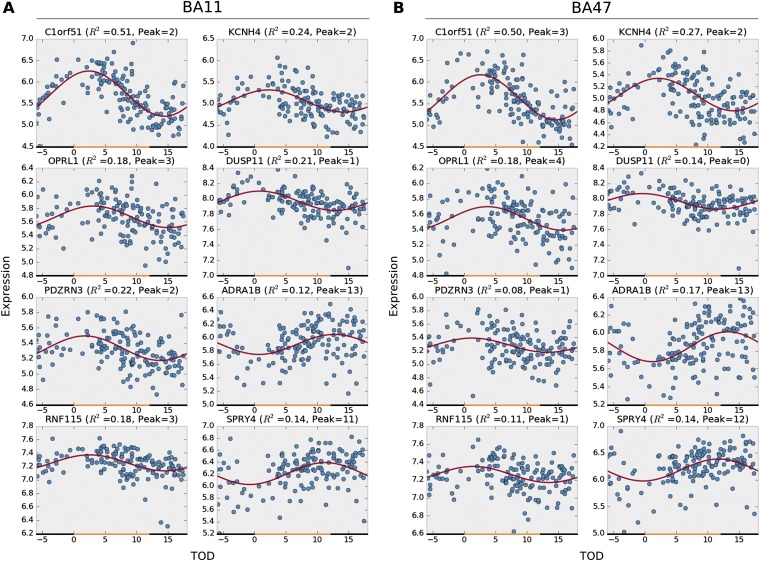
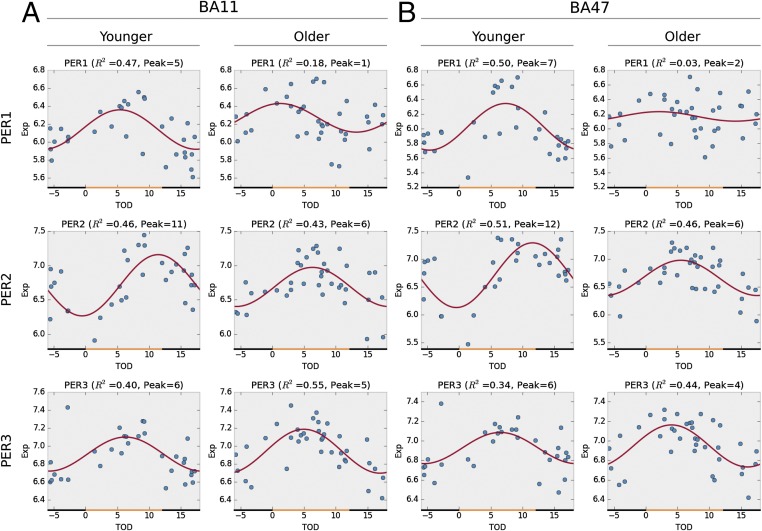
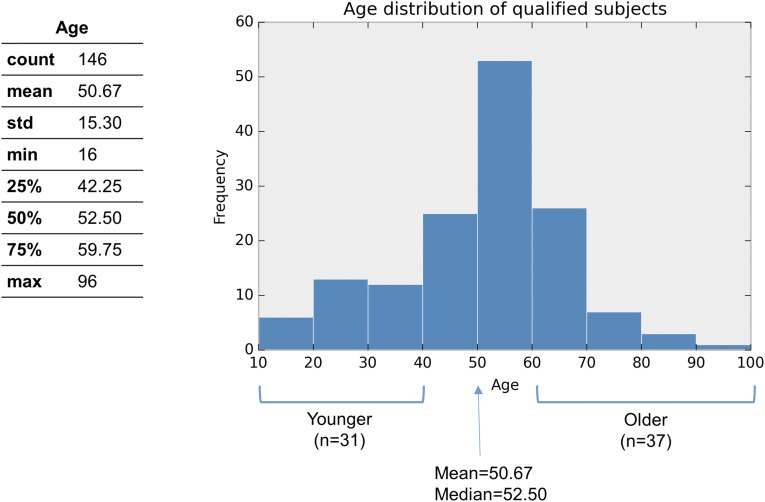
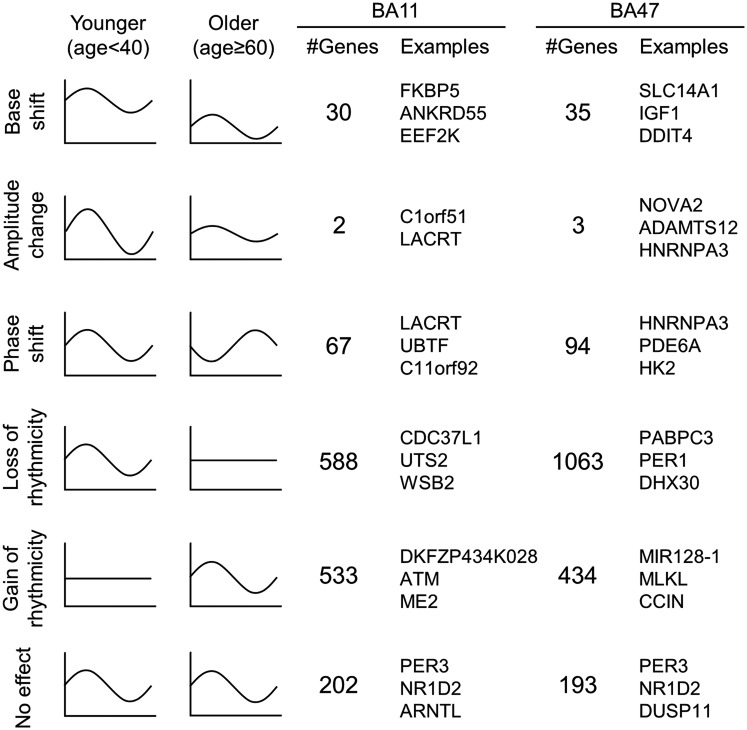
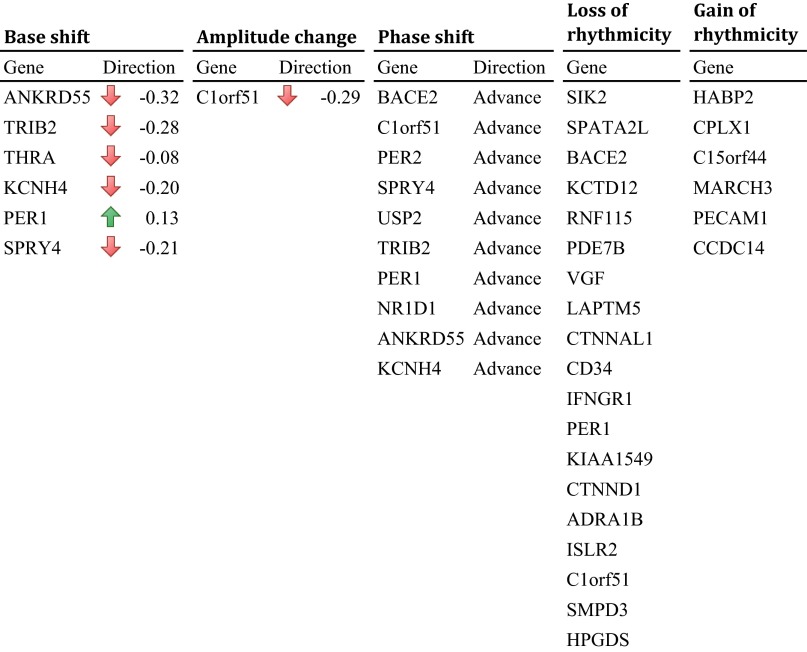
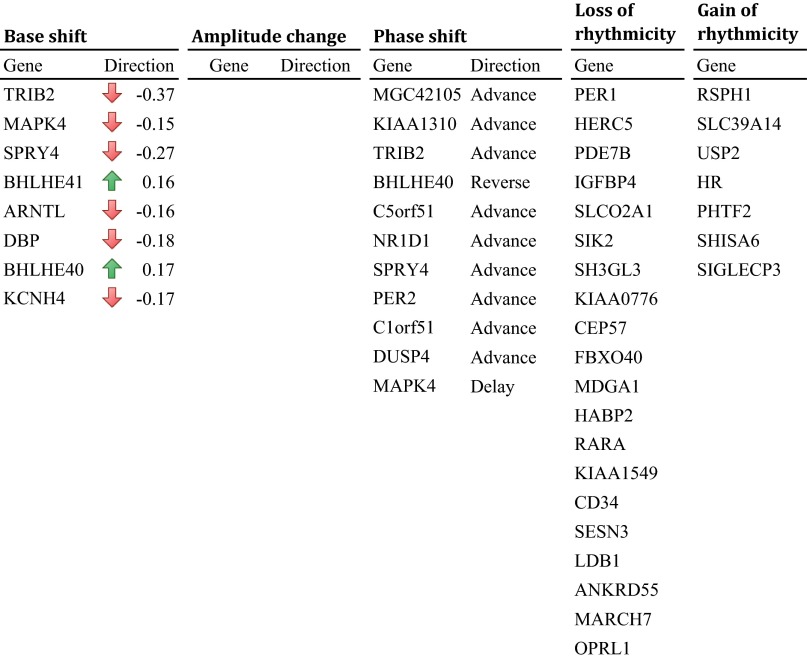
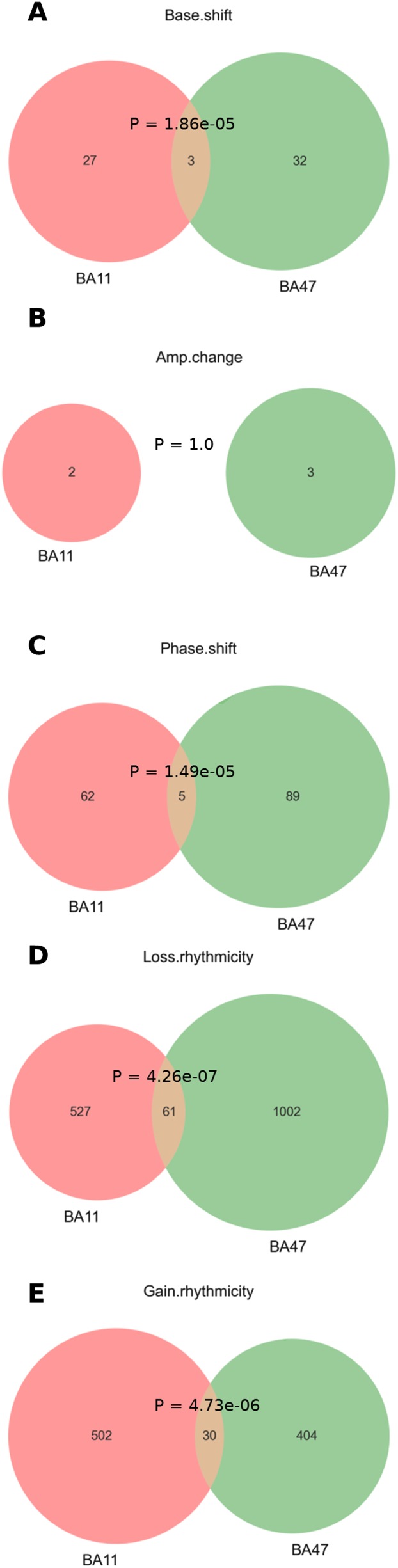
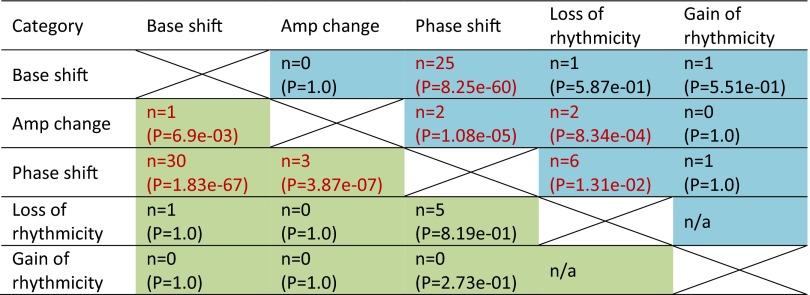
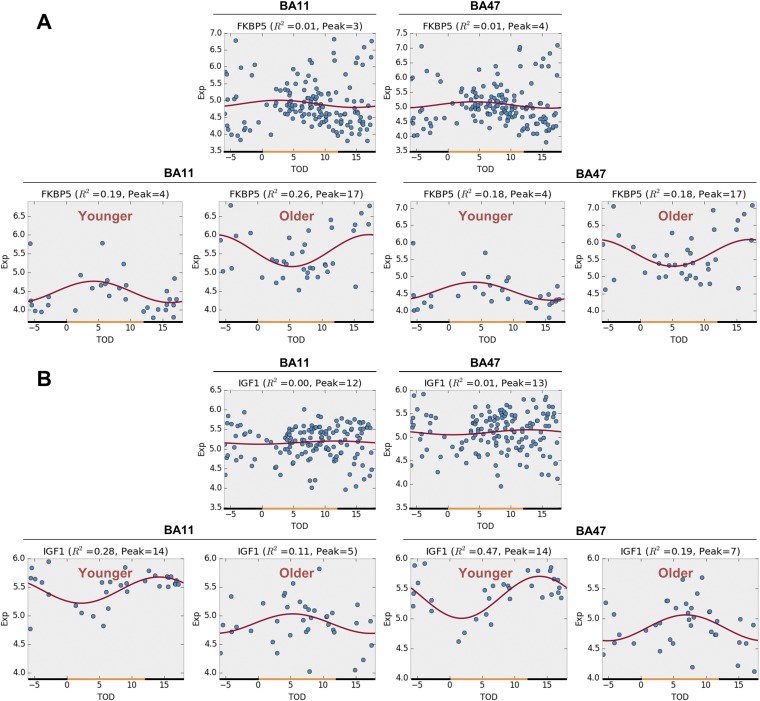
With aging, significant changes in circadian rhythms occur, including a shift in phase toward a "morning" chronotype and a loss of rhythmicity in circulating hormones. However, the effects of aging on molecular rhythms in the human brain have remained elusive. Here, we used a previously described time-of-death analysis to identify transcripts throughout the genome that have a significant circadian rhythm in expression in the human prefrontal cortex [Brodmann's area 11 (BA11) and BA47]. Expression levels were determined by microarray analysis in 146 individuals. Rhythmicity in expression was found in ∼ 10% of detected transcripts (P < 0.05). Using a metaanalysis across the two brain areas, we identified a core set of 235 genes (q < 0.05) with significant circadian rhythms of expression. These 235 genes showed 92% concordance in the phase of expression between the two areas. In addition to the canonical core circadian genes, a number of other genes were found to exhibit rhythmic expression in the brain. Notably, we identified more than 1,000 genes (1,186 in BA11; 1,591 in BA47) that exhibited age-dependent rhythmicity or alterations in rhythmicity patterns with aging. Interestingly, a set of transcripts gained rhythmicity in older individuals, which may represent a compensatory mechanism due to a loss of canonical clock function. Thus, we confirm that rhythmic gene expression can be reliably measured in human brain and identified for the first time (to our knowledge) significant changes in molecular rhythms with aging that may contribute to altered cognition, sleep, and mood in later life.
Keywords: aging; circadian rhythms; gene expression; postmortem; prefrontal cortex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Foster RG, Kreitzman L. The rhythms of life: What your body clock means to you! Exp Physiol. 2014;99(4):599–606. - PubMed
-
- Reppert SM, Weaver DR. Molecular analysis of mammalian circadian rhythms. Annu Rev Physiol. 2001;63:647–676. - PubMed
-
- Hogenesch JB, Panda S, Kay S, Takahashi JS. Circadian transcriptional output in the SCN and liver of the mouse. Novartis Found Symp. 2003;253:171–180; discussion 52–55, 102–109, 180–183 passim. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
