

p53 genes function to restrain mobile elements
- PMID: 26701264
- PMCID: PMC4701979
- DOI: 10.1101/gad.266098.115
p53 genes function to restrain mobile elements
Abstract
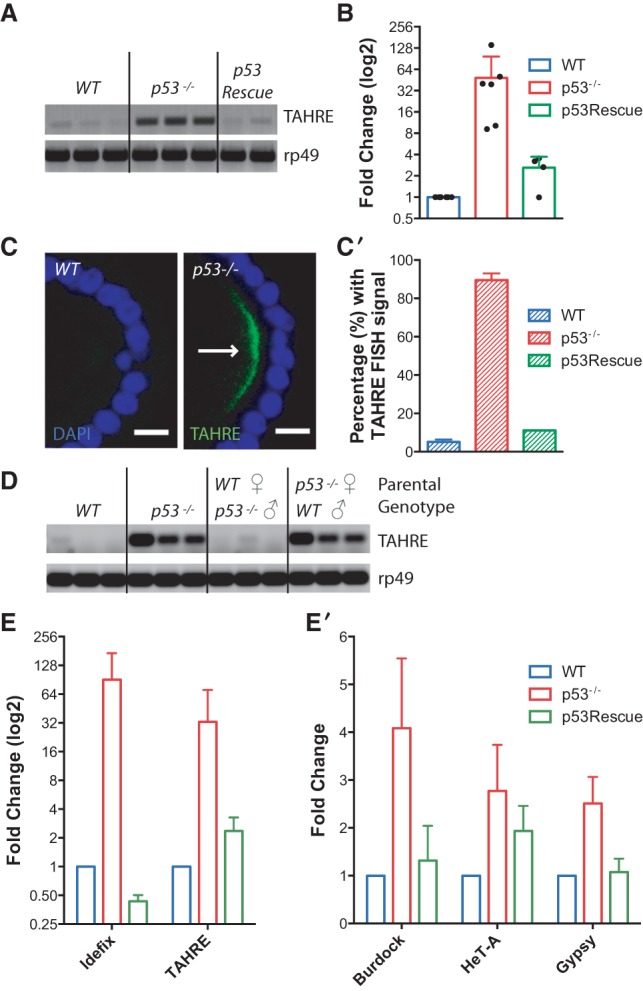
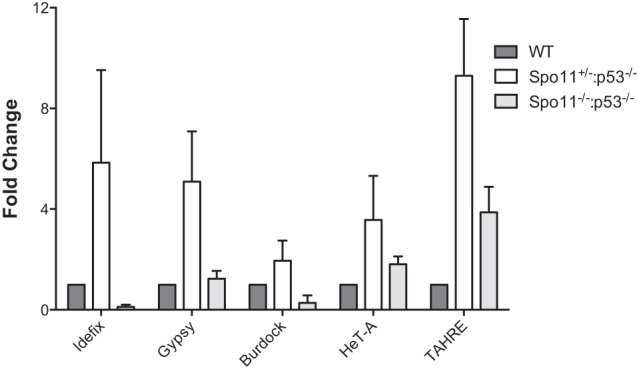
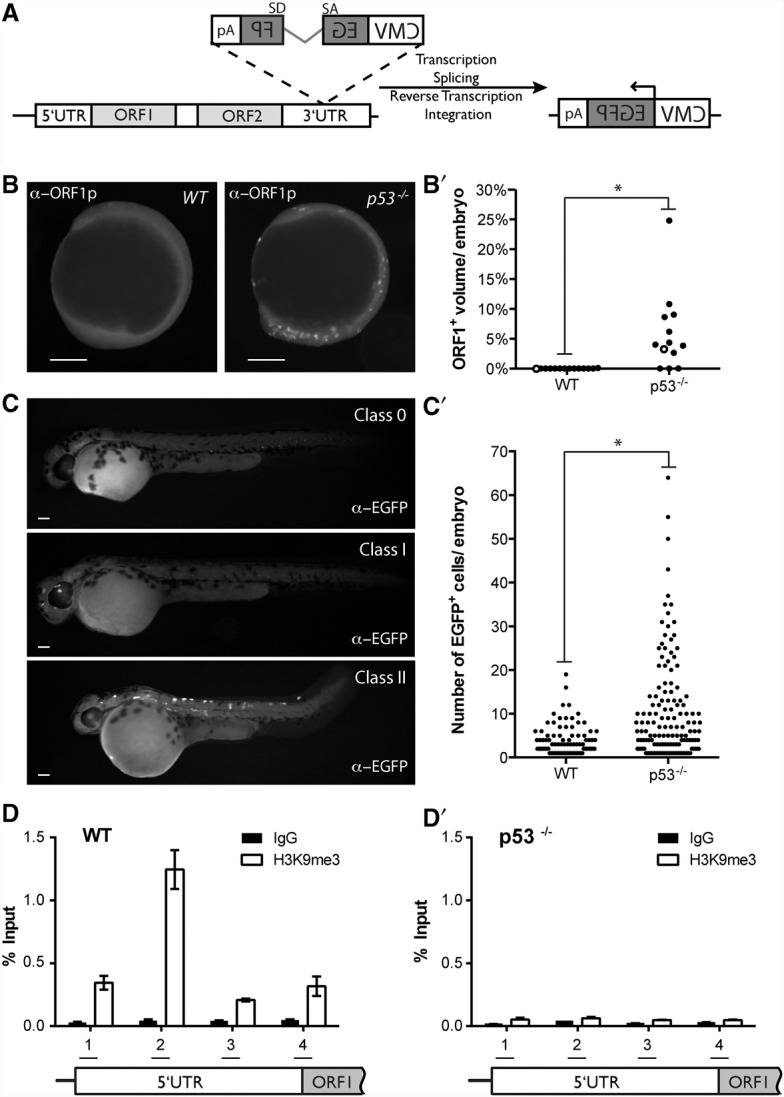
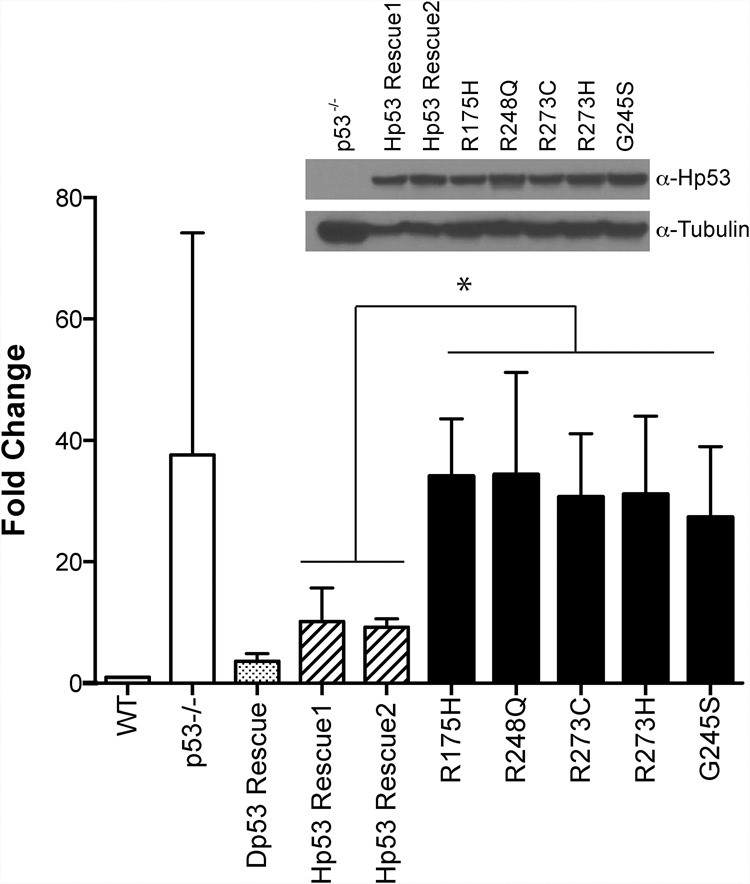
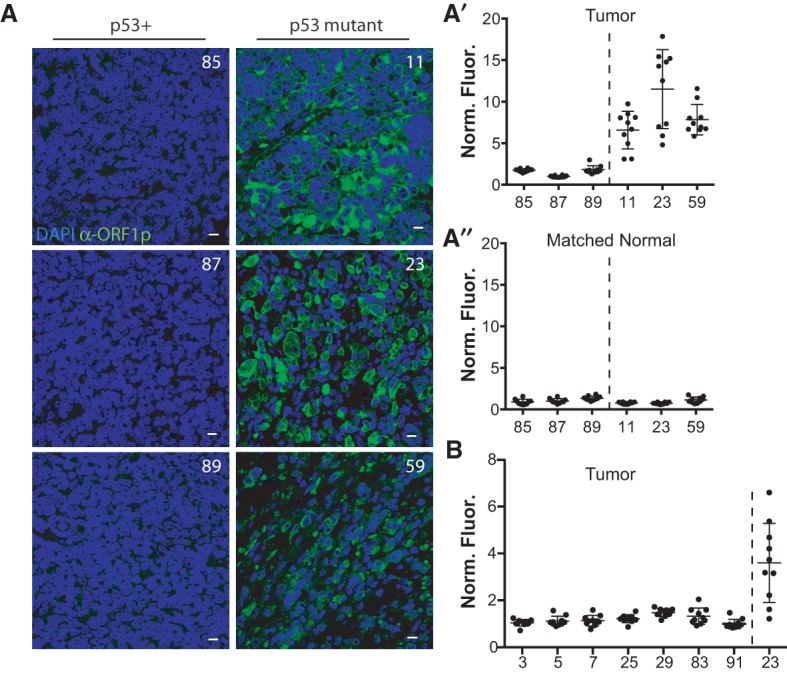
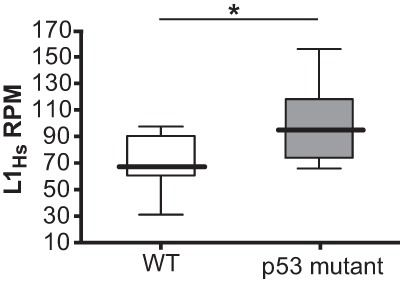
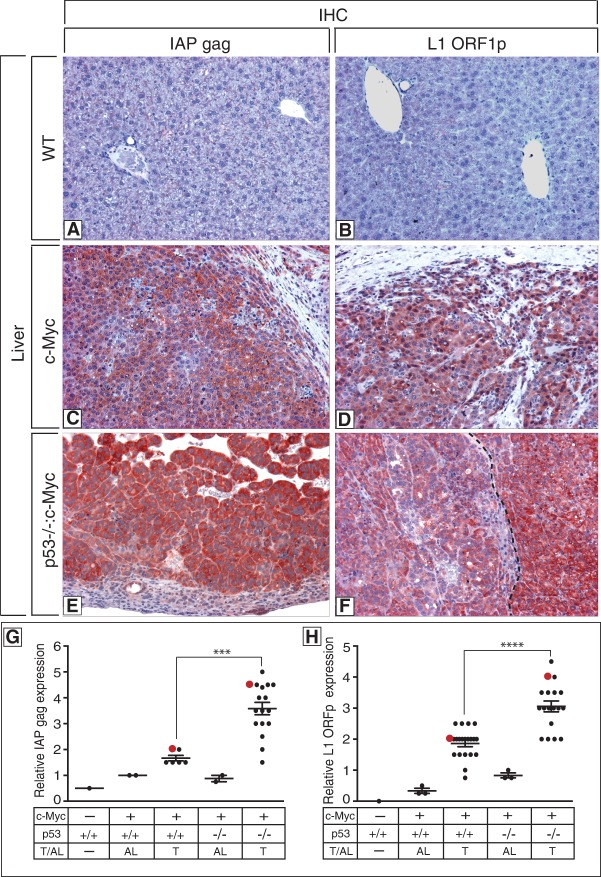
Throughout the animal kingdom, p53 genes govern stress response networks by specifying adaptive transcriptional responses. The human member of this gene family is mutated in most cancers, but precisely how p53 functions to mediate tumor suppression is not well understood. Using Drosophila and zebrafish models, we show that p53 restricts retrotransposon activity and genetically interacts with components of the piRNA (piwi-interacting RNA) pathway. Furthermore, transposon eruptions occurring in the p53(-) germline were incited by meiotic recombination, and transcripts produced from these mobile elements accumulated in the germ plasm. In gene complementation studies, normal human p53 alleles suppressed transposons, but mutant p53 alleles from cancer patients could not. Consistent with these observations, we also found patterns of unrestrained retrotransposons in p53-driven mouse and human cancers. Furthermore, p53 status correlated with repressive chromatin marks in the 5' sequence of a synthetic LINE-1 element. Together, these observations indicate that ancestral functions of p53 operate through conserved mechanisms to contain retrotransposons. Since human p53 mutants are disabled for this activity, our findings raise the possibility that p53 mitigates oncogenic disease in part by restricting transposon mobility.
Keywords: Drosophila; human cancers; mouse cancer models; p53; piRNAs; retrotransposons; zebrafish.
© 2016 Wylie et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Aravin AA, Hannon GJ, Brennecke J. 2007. The Piwi–piRNA pathway provides an adaptive defense in the transposon arms race. Science 318: 761–764. - PubMed
-
- Bogdanovic O, Fernández-Miñán A, Tena JJ, de la Calle-Mustienes E, Gómez-Skarmeta JL. 2013. The developmental epigenomics toolbox: ChIP-seq and MethylCap-seq profiling of early zebrafish embryos. Methods 62: 207–215. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA135731/CA/NCI NIH HHS/United States
- T32 DK007307/DK/NIDDK NIH HHS/United States
- R01GM072124/GM/NIGMS NIH HHS/United States
- T32 5T32DK007307/DK/NIDDK NIH HHS/United States
- R01 GM115682/GM/NIGMS NIH HHS/United States
- R01CA135731/CA/NCI NIH HHS/United States
- R01 GM060518/GM/NIGMS NIH HHS/United States
- R01GM115682/GM/NIGMS NIH HHS/United States
- F31 GM108472/GM/NIGMS NIH HHS/United States
- T32 GM083831/GM/NIGMS NIH HHS/United States
- P30 CA142543/CA/NCI NIH HHS/United States
- 1F31GM108472-01/GM/NIGMS NIH HHS/United States
- Howard Hughes Medical Institute/United States
- F31 CA189691/CA/NCI NIH HHS/United States
- 1F31CA189691-01/CA/NCI NIH HHS/United States
- GM060518/GM/NIGMS NIH HHS/United States
- R01 GM072124/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous