

Over-expression of Plk4 induces centrosome amplification, loss of primary cilia and associated tissue hyperplasia in the mouse
- PMID: 26701933
- PMCID: PMC4703062
- DOI: 10.1098/rsob.150209
Over-expression of Plk4 induces centrosome amplification, loss of primary cilia and associated tissue hyperplasia in the mouse
Abstract
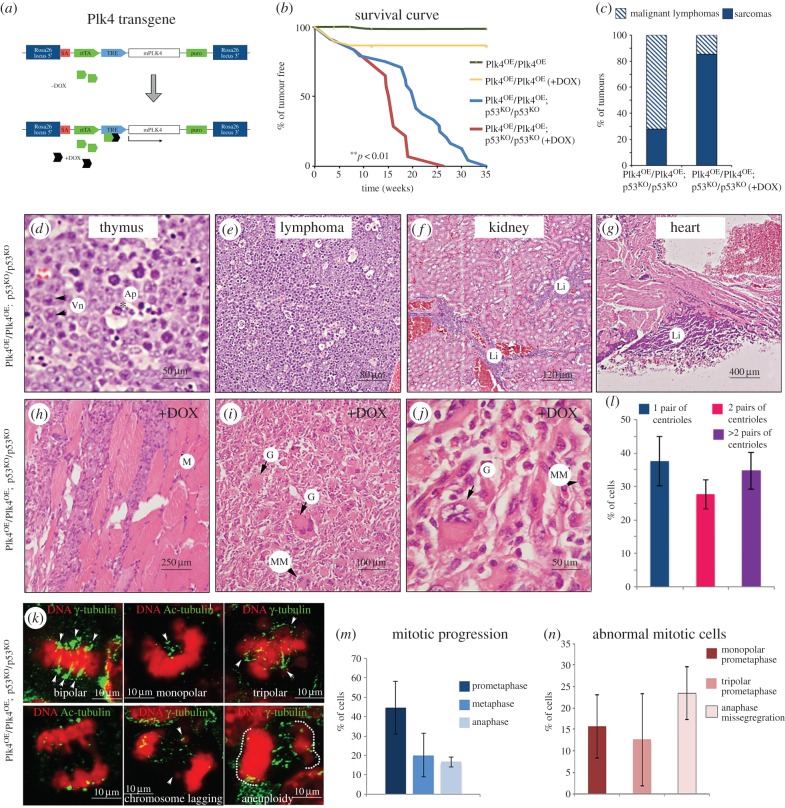
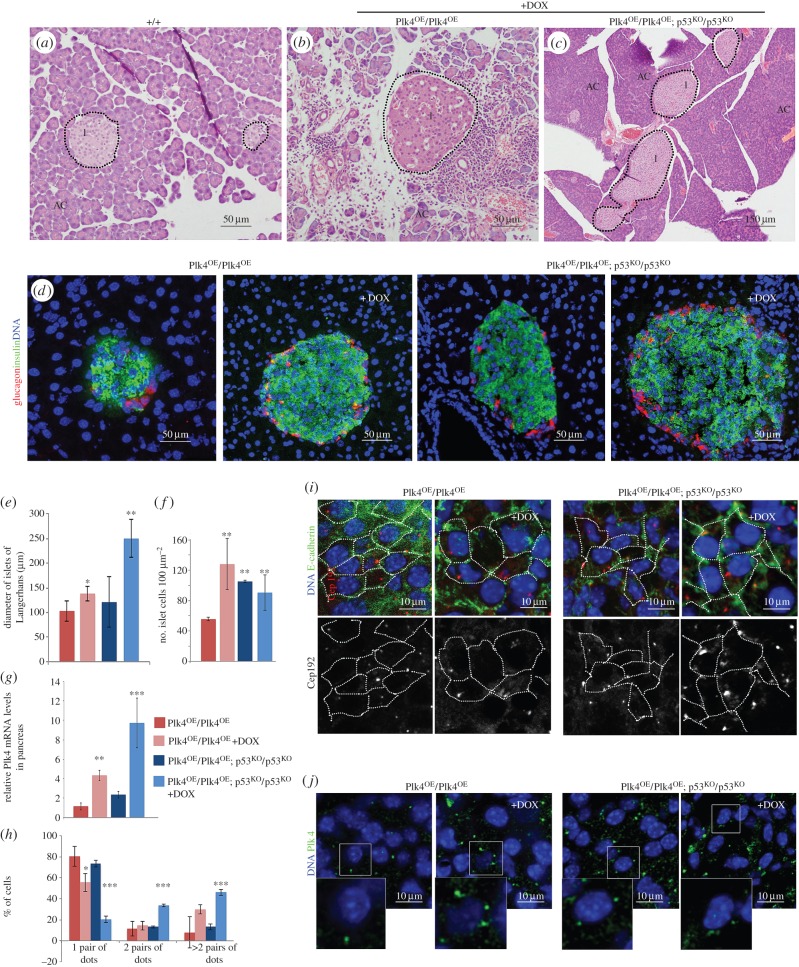
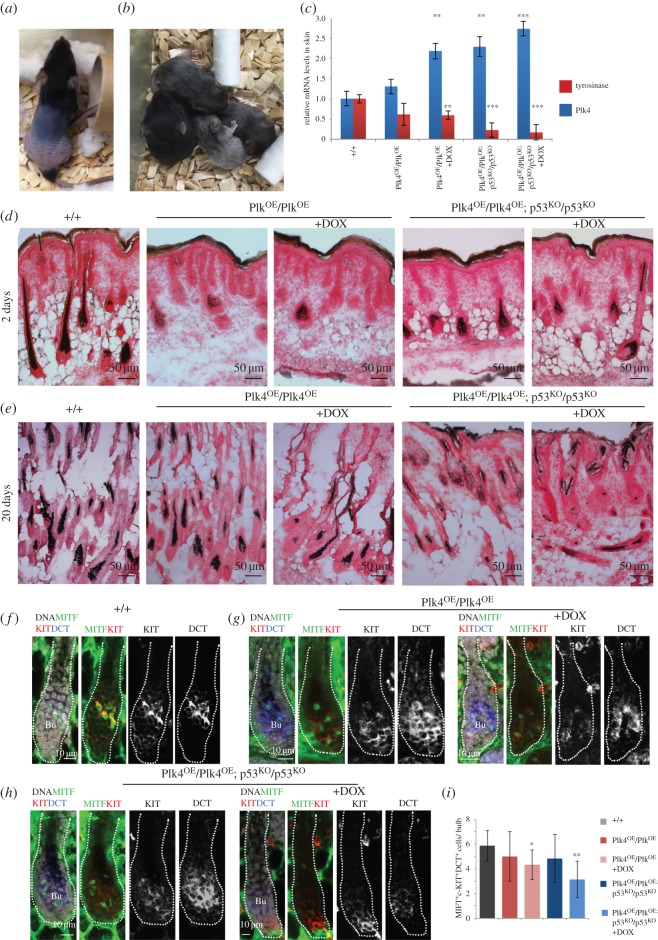
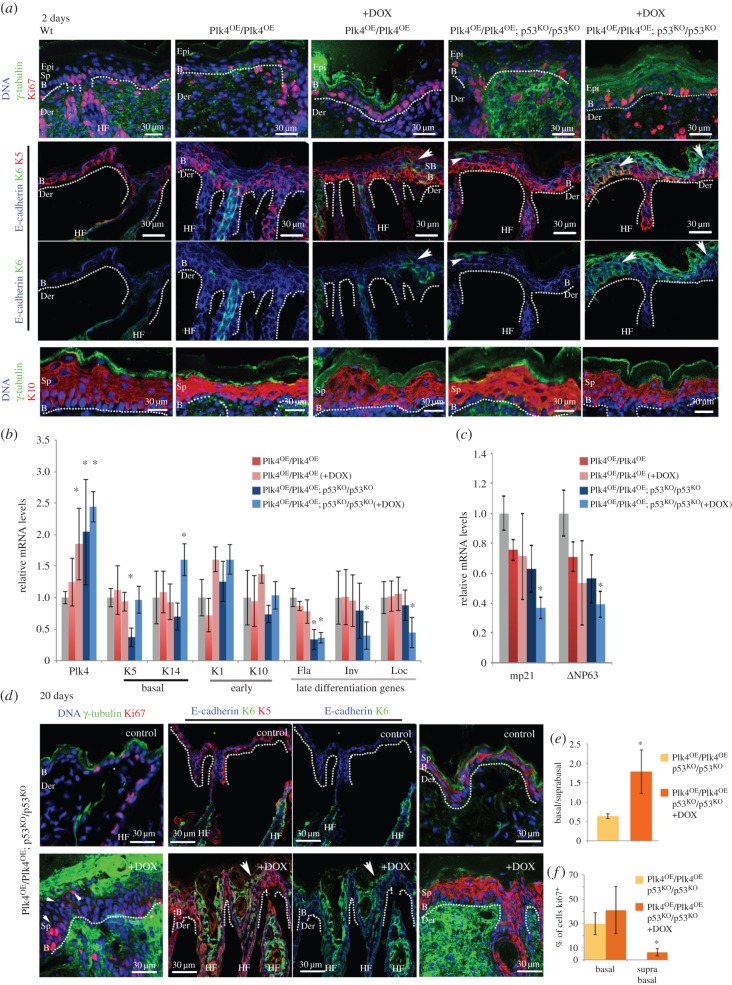
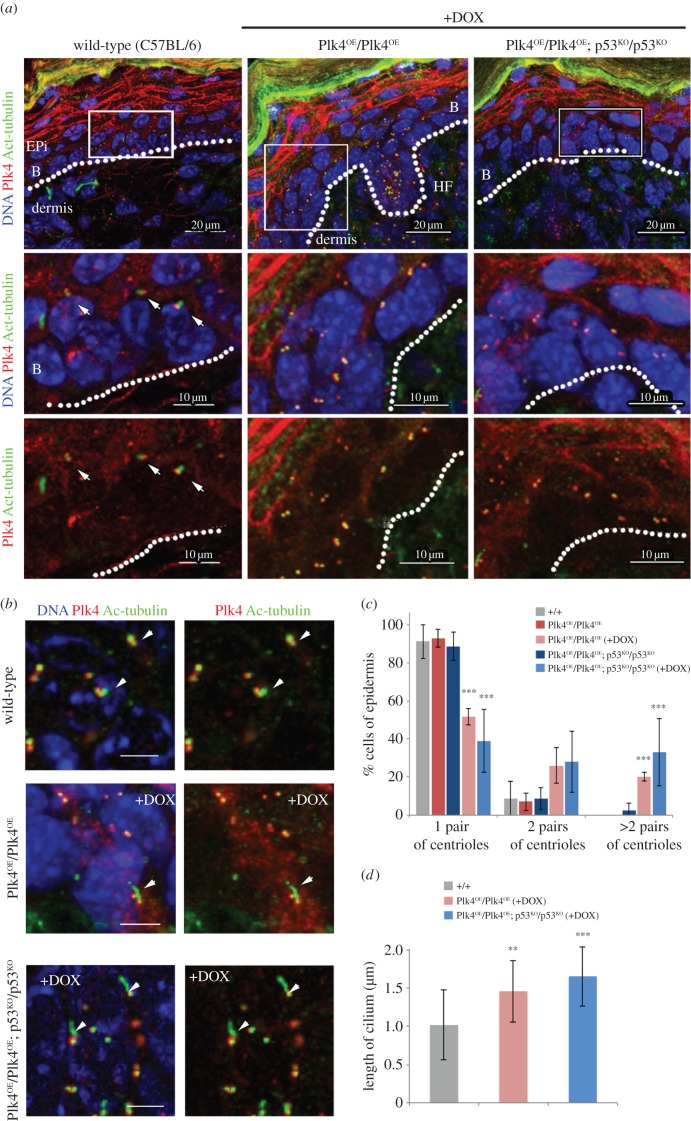
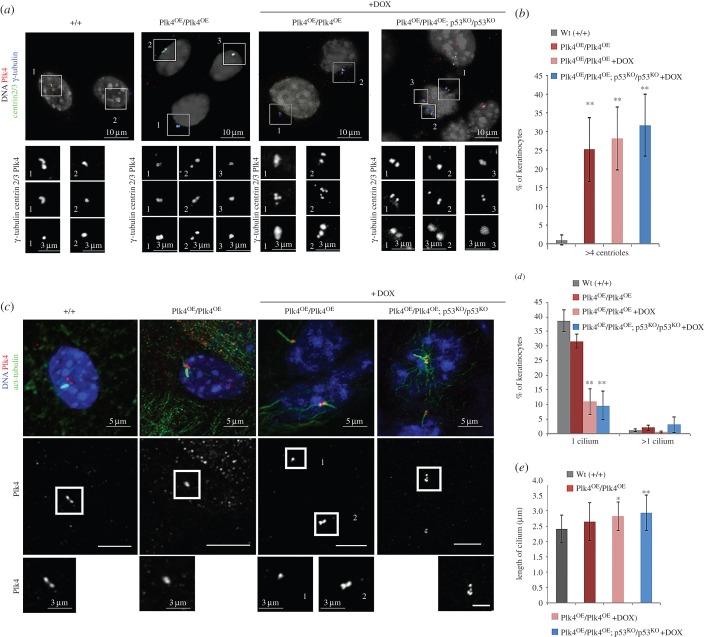
To address the long-known relationship between supernumerary centrosomes and cancer, we have generated a transgenic mouse that permits inducible expression of the master regulator of centriole duplication, Polo-like-kinase-4 (Plk4). Over-expression of Plk4 from this transgene advances the onset of tumour formation that occurs in the absence of the tumour suppressor p53. Plk4 over-expression also leads to hyperproliferation of cells in the pancreas and skin that is enhanced in a p53 null background. Pancreatic islets become enlarged following Plk4 over-expression as a result of equal expansion of α- and β-cells, which exhibit centrosome amplification. Mice overexpressing Plk4 develop grey hair due to a loss of differentiated melanocytes and bald patches of skin associated with a thickening of the epidermis. This reflects an increase in proliferating cells expressing keratin 5 in the basal epidermal layer and the expansion of these cells into suprabasal layers. Such cells also express keratin 6, a marker for hyperplasia. This is paralleled by a decreased expression of later differentiation markers, involucrin, filaggrin and loricrin. Proliferating cells showed an increase in centrosome number and a loss of primary cilia, events that were mirrored in primary cultures of keratinocytes established from these animals. We discuss how repeated duplication of centrioles appears to prevent the formation of basal bodies leading to loss of primary cilia, disruption of signalling and thereby aberrant differentiation of cells within the epidermis. The absence of p53 permits cells with increased centrosomes to continue dividing, thus setting up a neoplastic state of error prone mitoses, a prerequisite for cancer development.
Keywords: Polo-like-kinase-4; centrosome amplification; pancreas; primary cilia; skin; tumour development.
© 2015 The Authors.
Figures
References
-
- Scheer U. 2014. Historical roots of centrosome research: discovery of Boveri's microscope slides in Würzburg. Phil. Trans. R. Soc. B 369, 20130469 (doi:10.1098/rstb.2013.0469) - DOI - PMC - PubMed
-
- Krämer A, Neben K, Ho AD. 2005. Centrosome aberrations in hematological malignancies. Cell Biol. Int. 29, 375–383. (doi:10.1016/j.cellbi.2005.03.004) - DOI - PubMed
-
- Lingle WL, Lutz WH, Ingle JN, Maihle NJ, Salisbury JL. 1998. Centrosome hypertrophy in human breast tumors: Implications for genomic stability and cell polarity. Proc. Natl Acad. Sci. USA 95, 2950–2955. (doi:10.1073/pnas.95.6.2950) - DOI - PMC - PubMed
-
- Pihan GA, Purohit A, Wallace J, Knecht H, Woda B, Quesenberry P, Doxsey SJ. 1998. Centrosome defects and genetic instability in malignant tumors. Cancer Res. 58, 3974–3985. - PubMed
-
- Hsu L-C, Kapali M, DeLoia JA, Gallion HH. 2005. Centrosome abnormalities in ovarian cancer. Int. J. Cancer 113, 746–751. (doi:10.1002/ijc.20633) - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous