

Functional and Structural Divergence in Human TRPV1 Channel Subunits by Oxidative Cysteine Modification
- PMID: 26702055
- PMCID: PMC4759194
- DOI: 10.1074/jbc.M115.700278
Functional and Structural Divergence in Human TRPV1 Channel Subunits by Oxidative Cysteine Modification
Abstract
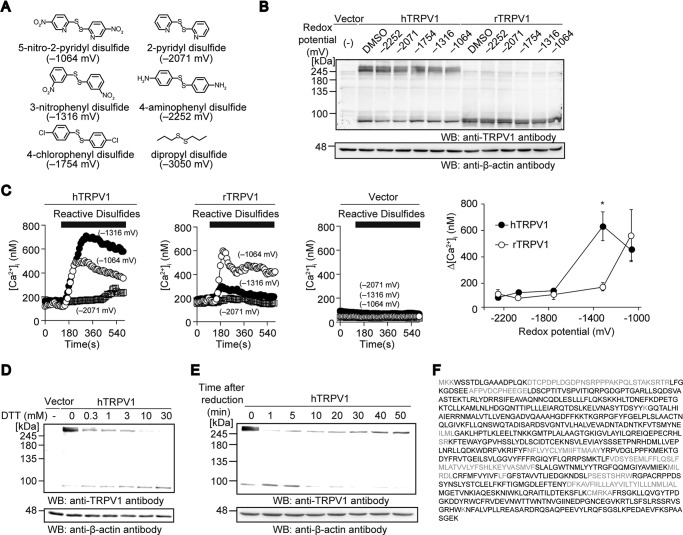
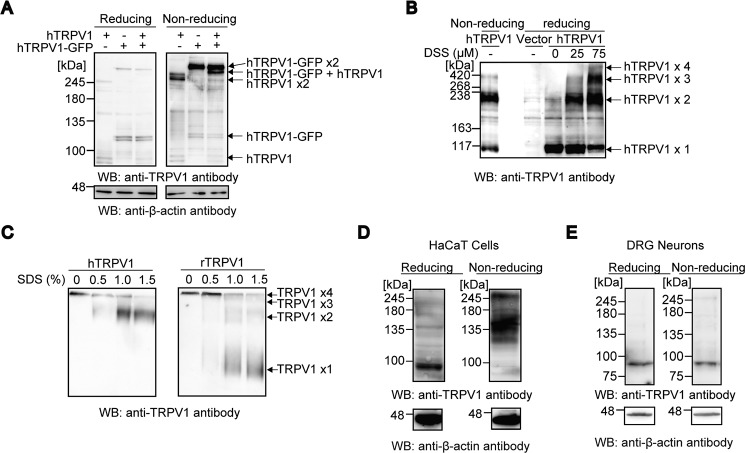
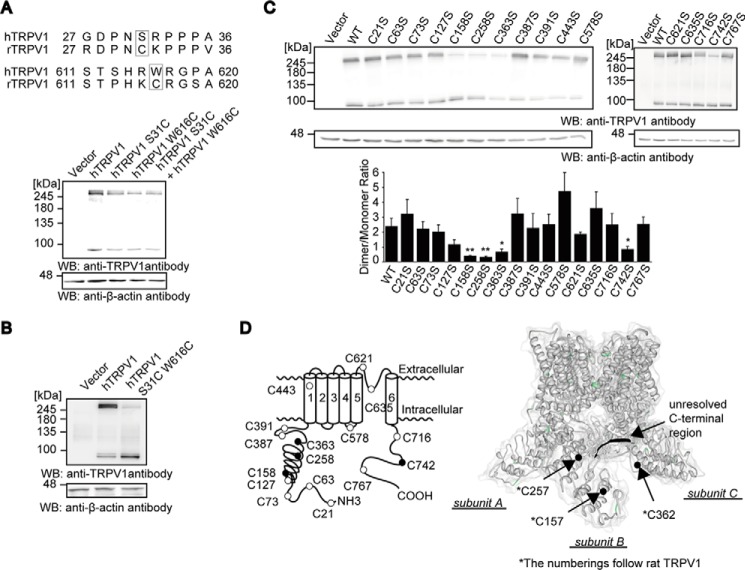
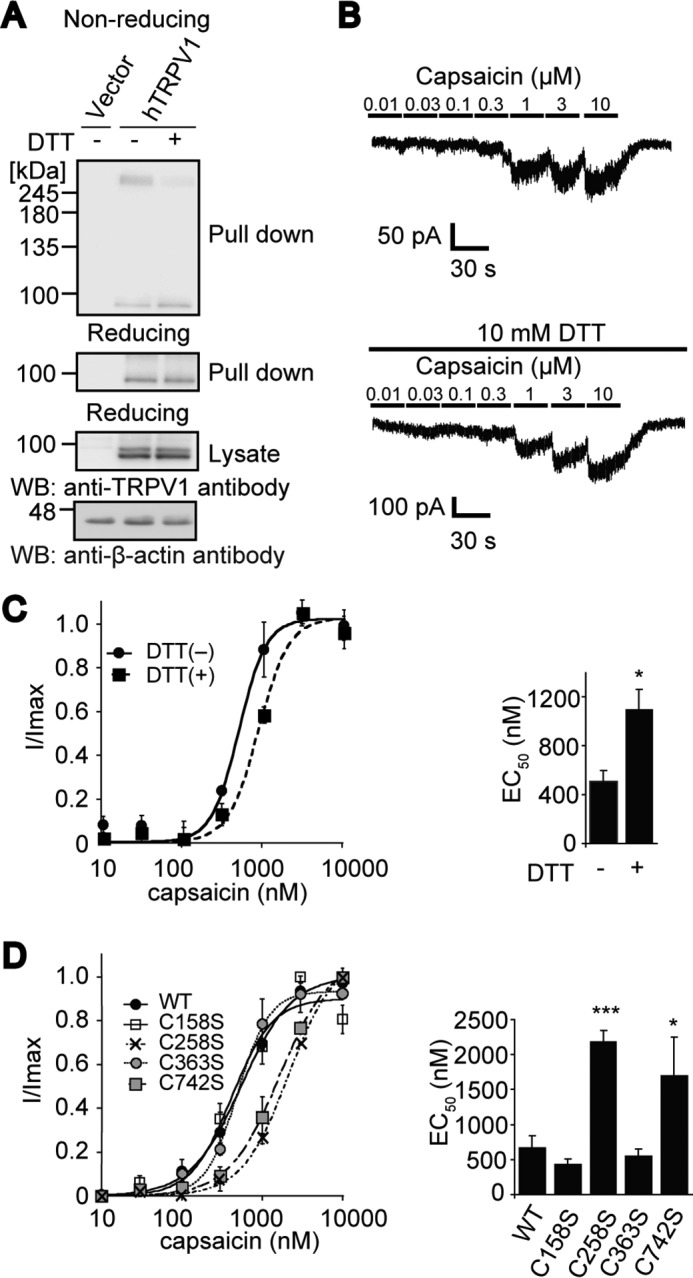
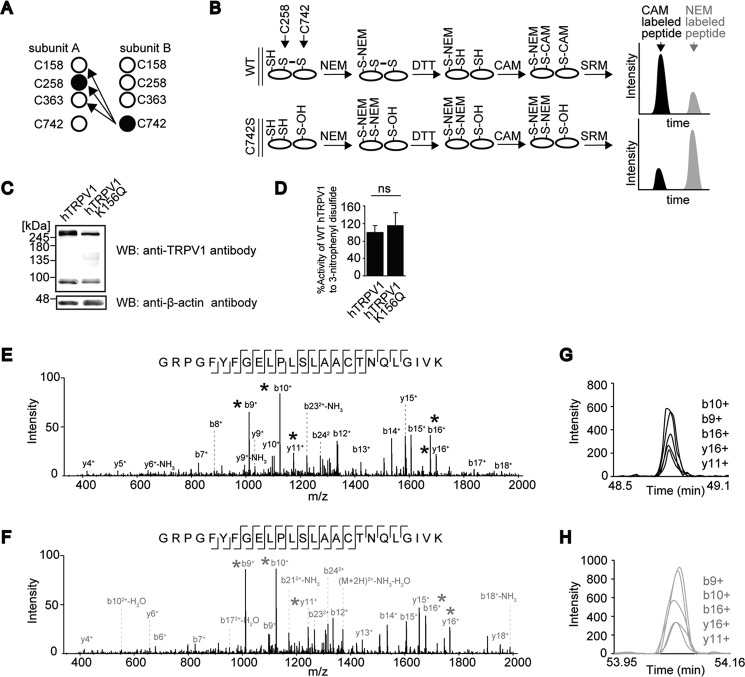
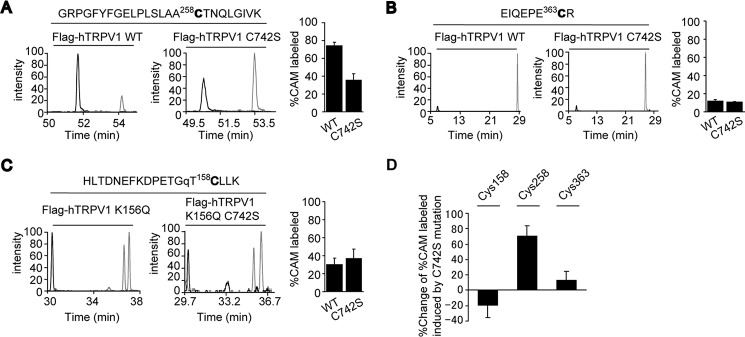
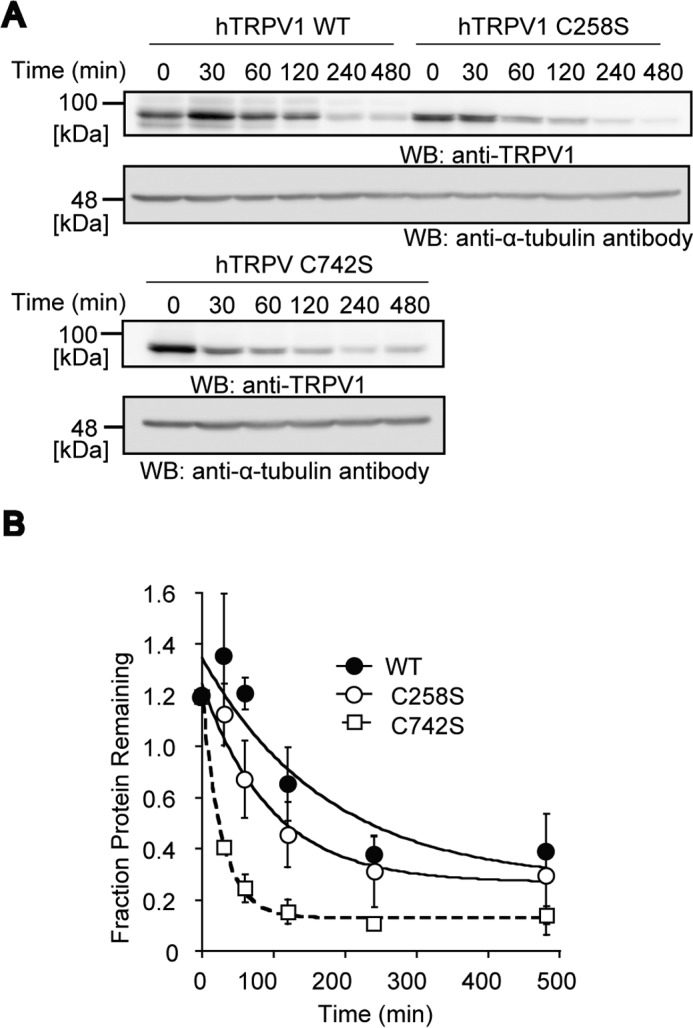
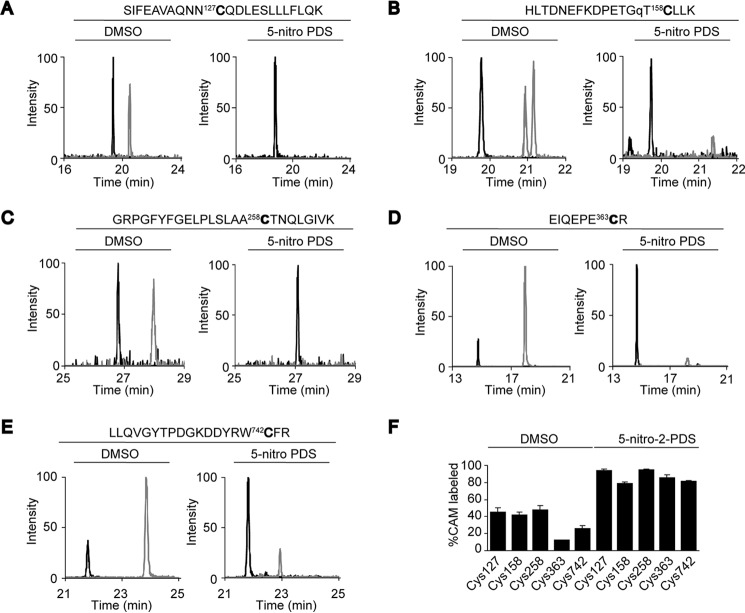
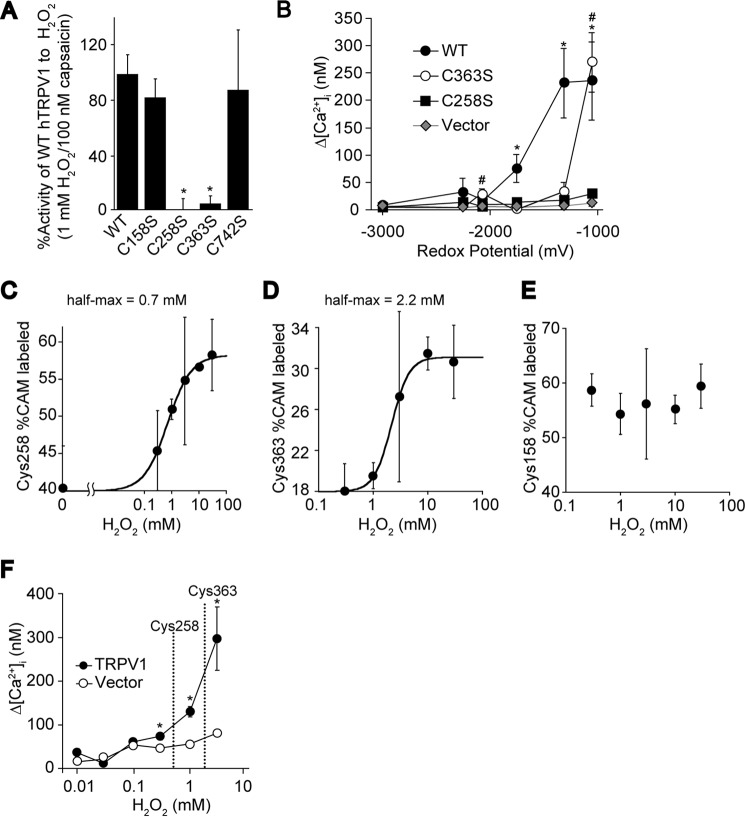
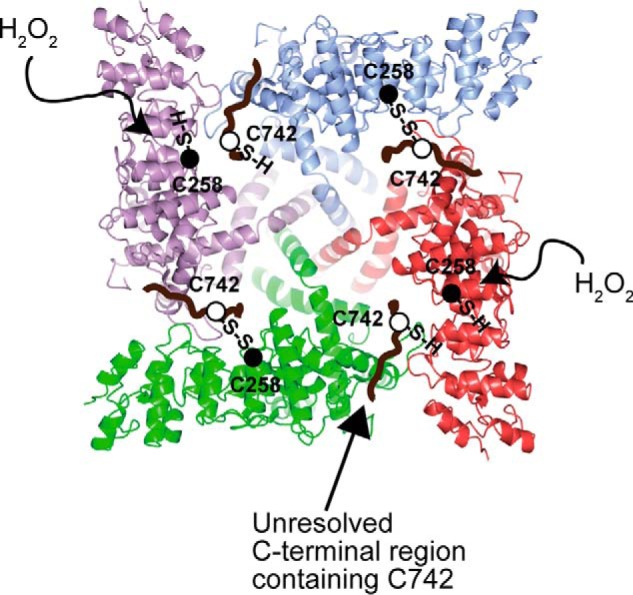
Transient receptor potential vanilloid 1 (TRPV1) channel is a tetrameric protein that acts as a sensor for noxious stimuli such as heat and for diverse inflammatory mediators such as oxidative stress to mediate nociception in a subset of sensory neurons. In TRPV1 oxidation sensing, cysteine (Cys) oxidation has been considered as the principle mechanism; however, its biochemical basis remains elusive. Here, we characterize the oxidative status of Cys residues in differential redox environments and propose a model of TRPV1 activation by oxidation. Through employing a combination of non-reducing SDS-PAGE, electrophysiology, and mass spectrometry we have identified the formation of subunit dimers carrying a stable intersubunit disulfide bond between Cys-258 and Cys-742 of human TRPV1 (hTRPV1). C258S and C742S hTRPV1 mutants have a decreased protein half-life, reflecting the role of the intersubunit disulfide bond in supporting channel stability. Interestingly, the C258S hTRPV1 mutant shows an abolished response to oxidants. Mass spectrometric analysis of Cys residues of hTRPV1 treated with hydrogen peroxide shows that Cys-258 is highly sensitive to oxidation. Our results suggest that Cys-258 residues are heterogeneously modified in the hTRPV1 tetrameric complex and comprise Cys-258 with free thiol for oxidation sensing and Cys-258, which is involved in the disulfide bond for assisting subunit dimerization. Thus, the hTRPV1 channel has a heterogeneous subunit composition in terms of both redox status and function.
Keywords: dimerization; disulfide bond; pain; post-translational modification (PTM); reactive oxygen species (ROS); redox regulation; transient receptor potential channels (TRP channels).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Clapham D. E. (2003) TRP channels as cellular sensors. Nature 426, 517–524 - PubMed
-
- Clapham D. E., Julius D., Montell C., and Schultz G. (2005) International Union of Pharmacology. XLIX. Nomenclature and structure-function relationships of transient receptor potential channels. Pharmacol. Rev. 57, 427–450 - PubMed
-
- Kozai D., Ogawa N., and Mori Y. (2014) Redox regulation of transient receptor potential channels. Antioxid. Redox. Signal. 21, 971–986 - PubMed
-
- Susankova K., Tousova K., Vyklicky L., Teisinger J., and Vlachova V. (2006) Reducing and oxidizing agents sensitize heat-activated vanilloid receptor (TRPV1) current. Mol. Pharmacol. 70, 383–394 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
